|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Поперечно – полосатые мышцы.
Поперечно-полосатые мышцы осуществляют передвижение тела в пространстве, перемещение частей тела относительно друг друга, поддержание позы, выработку тепла. Они обладают тремя важнейшими свойствами: 1) возбудимостью, т.е. способностью отвечать на действие раздражителя генерацией ПД; 2) проводимостью, способностью к проведению ПД вдоль всего мышечного волокна; 3) сократимостью, способностью сокращаться или изменять напряжение при возбуждении. В организме возбуждение и сокращение мышц вызывается нервными импульсами. Иннервация поперечно-полосатых мышечных волокон позвоночных осуществляется из мотонейронов. Одно нервное волокно имеет синаптические контакты с целой группой мышечных волокон. Такая группа называется нейромоторной единицей. Мышечные волокна, входящие в состав одной нейромоторной единицы возбуждаются синхронно. Волокна, входящие в состав разных двигательных единиц могут возбуждаться асинхронно. Это обусловлено тем, что сила раздражения является пороговой не для всех нервных волокон. Если в сократительном ответе приняли участие все моторные единицы, мышца сокращается с максимальной ампдитудой. В эксперименте применяют электрическое раздражение. Непосредственное раздражение самой мышцы называют прямым раздражением. Раздражение двигательного нерва, иннервирующего мышцу, называют непрямое раздражение. Возбудимость мышечной ткани ниже, чем нервной, и электрическое раздражение непосредственно мышцы еще не обеспечивает прямого раздражения: ток, распространяясь по мышечной ткани, действует в первую очередь на находящиеся в ней окончания двигательных нервов и возбуждает их, что ведет к сокращению мышц. В эксперименте эффект чисто прямого раздражения достигается блокадой нервно – мышечного синапса ядом кураре, или электрическим раздражением с помощью микроэлектрода, введенного внутрь мышечного волокна. Электрическую активность целой мышцы при возбуждении можно зарегистрировать при помощи приложенных к мышце электродов и дальнейшего усиления отводимых потенциалов. Методика регистрации электрической активности мышц получила название электромиография. Электромиография находит широкое применение в физиологии спорта и медицине для оценки состояния двигательного аппарата, Различают два вида мышечного сокращения - изотоническое и изометрическое. Изотоническим называют такое сокращение, при котором происходит укорочение длины мышцы без изменения напряжения. Изометрическое сокращение сопровождается изменением напряжения без изменения длины. Естественные сокращения никогда не бывают чисто изотоническими, так поднимая постоянный груз, мышца изменяет свое напряжение вследствие реальной нагрузки. К целостному организму применяется иная классификация типов сокращения: выделяют изометрическое сокращение, концентрическое, при котором мышца укорачивается и эксцентрическое, совершаемое в условиях удлинения мышцы. Возбудимость и возбуждение мышечных волокон. Механизм генерации потенциалов в скелетных мышечных волокнах принципиально не отличается от такового в нервных волокнах. Величина потенциала покоя (ПП) в скелетных мышечных волокнах составляет = -90 мВ, критический потенциал – 50 мВ (как и в нервных волокнах), амплитуда потенциала действия (ПД) = 120 -130 мВ, длительность ПД = 2-3 мсек (для мышц конечностей), скорость распространения потенциала действия = 3 – 5 м/сек. В скелетных мышцах человека и теплокровных животных различают быстрые двигательные единицы, длительность сокращения которых составляет 10 – 30 мсек и медленные - с длительностью 100 мсек. Медленные двигательные волокна содержат меньше мышечных волокон и развивают меньшую силу, устойчивы к утомлению, их мотонейроны более возбудимы. При тоническом напряжении, связанном с поддержанием позы активируются только низкопороговые, медленные, устойчивые к утомлению двигательные единицы. В естественных условиях мышца редко бывает расслабленной. Наблюдается небольшая, так называемая, позная активность (позный тонус). Одиночное мышечное сокращение. Одииночный стимул вызывает одиночное сокращение мышцы. Различают две основные фазы мышечного сокращения: фазу укорочения и фазу расслабления. От момента нанесения раздражения до выраженной фазы сокращения отмечается очень короткий скрытый (латентный) период. Было установлено, что сокращение мышечного волокна начинается уже во время восходящей фазы ПД и начало его совпадает с моментом, когда ПД достигает пороговой для мышечного ответа величины (40 мВ). Длительность сокращения в десятки раз превышает продолжительность ПД. Потенциал действия, пройдя вдоль всего волокна, заканчивается, а мышечное сокращение достигает максимального напряжения или укорочения. Амплитуда сокращения одиночного мышечного волокна подчиняется закону “ все или ничего”. Сокращение же целой мышцы зависит от силы раздражения. При увеличении силы раздражения число возбужденных волокон возрастает. Мышечное сокращение увеличивается до тех пор, пока все волокна не оказываются сокращенными.
Рис. 28. Кривые двух одиночных сокращений изолированного мышечного волокна. Оптическая регистрация. М — миограмма; Р —-отметка раздражения; В — отметка времени 0, 2 с (по Ф. Н. Серкову).
Рис. 29. Суммация мышечных сокращений в ответ на 2 стимула. Отметка времени 20 мс. Объяснение в тексте.
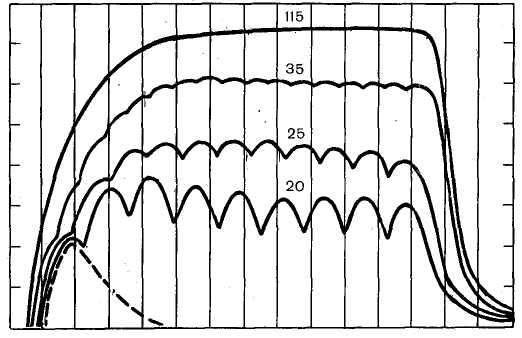
Рис. 30. Сокращения икроножной мышцы при разной частоте раздражения (по Е. Е. Жукову). Одиночное сокращение показано пунктиром. Цифры над кривыми — частоты стимулов. При частотах 20—35 в секунду — зубчатый тетанус, при частоте 1 15 в секунду — гладкий тетанус.
Рис.19. Одиночное и тетаническое сокращение мышцы.
Суммация сокращения. При ритмическом раздражении мышцы с частотой, когда происходит суммация эффектов, наступает сильное и длительное сокращение мышцы - тетаническое сокращение (или тетанус). Тетаническое сокращение - это результат суммации сократительных ответов, а не электрических реакций (ПД не суммируется) и его частота соответствует частоте ритмического раздражения, вызвавшего тетанус. Для возникновения тетануса необходимо, чтобы интервал между раздражениями был длиннее рефрактерного периода и короче всей длительности сократительного ответа. При относительно малой частоте наблюдается зубчатый тетанус. В этом случае второе раздражение поступает в фазу развития мышечного расслабления и вершина второго сократительного ответа будет отделена от вершины первого. При большой частоте раздражения возникает гладкий тетанус, так как очередное раздражение действует в момент развития мышечного сокращения и второе сокращение полностью сливается с первым (Рис.19). Причем амплитуда тетануса может быть больше величины максимального одиночного сокращения. Роль АТФ в механизме мышечного сокращения. Впервые В.А. Энгельгард и М. Н. Любимова в 19З9 году нашли, что мышечный белок миозин обладает свойствами АТФ-азы. Затем был открыт белок актин и показано, что он активирует АТФ-азную активность миозина. Для объяснения механизма мышечного сокращения используется теория скольжения нитей, независимо друг от друга в 1954 году предложенная двумя учеными – однофамильцами – А. Хаксли и Г. Хаксли. Согласно этой теории, сокращение мышцы происходит благодаря активному скольжению тонких актиновых нитей относительно толстых миозиновых. Процесс скольжения связан с взаимодействием активных центров белков миозиновых и актиновых нитей с образованием, так называемых, поперечных мостиков (поперечные мостики – это не что иное, как головки миозиновых нитей, изменяющие свое положение при связывании с актиновой нитью) в присутствии ионов кальция и АТФ. В состоянии покоя, основная часть ионов кальция хранится в саркоплазматическом ретикулуме скелетных мышечных волокон. Саркоплазматический ретикулум представляет собой замкнутую систему внутриклеточных трубочек и цистерн, окружающих каждую миофибриллу. Две важнейшие транспортные системы обеспечивают накопление в ретикулуме ионов кальция (секвестрация - захват их из миоплазмы) и освобождение кальция из ретикулума при возбуждении. В покое, в условиях низкой концентрации ионов Са++ в миоплазме, взаимодействию актиновых и миозиновых нитей препятствует белок тропонин, расположенный на актиновых нитях (Рис. 20). Как только концентрация Са++ в области миофибрилл повышается, тропонин связывает Са++ и изменяет свое расположение на актиновой нити, делая возможным взаимодействие актина с головкой миозина. Формирующиеся поперечные мостики перемещают актиновую нить относительно миозиновых всего на 10 нм. Это соответствует укорочению каждого саркомера лишь на 1%. Для дальнейшеего передвижение нити и соответственно сокращения волокна, необходимо, чтобы эти мостики разъединились и прикрепились к новым участкам актиновой нити. Такое разъединение мостиков осуществляется при расщеплении молекул АТФ АТФ – азой миозина, активизирующейся при связывании ионов кальция тропонином. В присутствии ионов кальция и АТФ в миоплазме этот процесс многократно повторяется: мостики повторно образуются и расходятся, в результате чего актиновые нити " скользят" и мышечное волокно укорачиватся. Для расслабления мышечного волокна необходима cеквестрация ионов Ca++ из миоплазмы в саркоплазматический ретикулум. Транспорт ионов Ca++ осуществляется против сил концентрационного градиента с использованием энергии АТФ.
Рис. 124. Схема, иллюстрирующая взаимоотношения между актином, тропомиозином и миозином при мышечном сокращении.
Таким образом, на основании выше изложенного, можно заключить, что энергия АТФ используется во время деятельности скелетной мышцы для трех основных процессов: 1. Работы натрий - калиевого насоса, обеспечивающего для поддержание постоянства градиента концентрации ионов Nа+ и К+ по обе стороны мембраны; 2. Процесса " скольжения" актиновых и миозиновых нитей, ведущего к укорочению миофибрилл; 3. Работы кальциевого насоса, необходимого для расслабления волокна. В соответствии с этим, ферменты, расщепляющие АТФ, локализованы в трех различных структурах мышечного волокна: клеточной мембране, миозиновых нитях и мембранах саркоплазматического ретикулума. Существуют два пути ресинтеза АТФ: быстрый, занимающий тысячные доли секунды, связанный с ферментативным переносом фосфатной группы от креатинфосфата на АДФ и более медленный, связанный с гликолитическими и окислительными процессами, протекающими в мышце, как в условиях покоя, так и особенно интенсивно во время деятельности. Во второй половине 19 века Гельмгольцем и В.Я. Данилевским было открыто образование тепла в мышечной ткани. С помощью высокочувствительных приборов Хиллу и сотр. удалось зарегистрировать образование тепла в покое и при мышечном сокращении. Первая фаза теплообразования начинается с момента возбуждения, непосредственно после нанесения раздражения, продолжается в течение всего сокращения, включая фазу расслабления (тепло активации, тепло укорочения, тепло расслабления). Первая фаза очень короткая, приблизительно, в 1000 раз короче второй и называется фазой начального теплообразования. Вторая фаза длится несколько минут и носит название запаздывающего, или восстановительного теплообразования. Она связана с химическими процессами, обеспечивающими ресинтез АТФ.
1.8.2. Гладкие мышцы. Гладкие мышцы в организме высших животных и человека находятся во внутренних органах, сосудах и коже. Гладкие мышцы образуют стенки полых органов: желудка, кишок, протоков пищеварительных желез, мочевого пузыря, желчного протока. Длительное тоническое сокращение гладких мышц, особенно выраженное в сфинктерах полых органов, препятствует выходу содержимого, обеспечивает накопление желчи в желчном пузыре, мочи в мочевом пузыре и т.д. Гладкие мышцы стенок кровеносных сосудов (артерий и артериол) регулируют величину просвета сосудов и тем самым уровень кровяного давления и кровоснабжения органов. Регуляция функции гладких мышц осуществляется гуморальными факторами и вегетативной нервной системой. К физиологическим особенностям гладких мышц относятся: 1) пластичность - способность после снятия груза оставаться растянутой без изменения напряжения; 2) образование функционального синцития, обеспечивающего передачу возбуждения с одной клетки на другую с вовлечением всей мышцы, несмотря на то, что двигательные нервные окончания расположены только на небольшом числе волокон гладких мышц (исключением являются реснитчатая мышца глаза и радиальная мышца радужной оболочки, каждое волокно которой имеет самостоятельную иннервацию, подобно волокнам скелетной мышцы); 3) автоматия - способность к спонтанной автоматической деятельности. Автоматия гладких мышц имеет миогенную природу и регулируется нервными элементами, которые находятся в стенках гладкомышечных органов. Адекватным раздражителем гладких мышц является их быстрое и сильное растяжение, в результате чего возникает деполяризация мембраны мышечного волокна и возникновение потенциала действия, способного к распространению. Потенциал покоя гладкомышечных волокон, обладающих автоматией, составляет 30 -70 мв, не обладающих автоматией - 60-70 мв. Амплитуда потенциала действия несколько ниже, чем в скелетных мышцах. Зарегистрированы два типа потенциаала действия - пикоподобные, длительностью от 5 до 80 мсек. и потенциалы с выраженным плато, продолжительностю 30-500 мсек. Деполяризация мембраны гладкомышечных волокон связана с активацией электровозбудимых Са++- каналов, отличающихся медленной кинетикой процессов активации и инактивации. Кальциевые каналы не чувствительны к тетродотоксину, но блокируются изоптином (верапамилом). Этот препарат используется в медицинской практике для устранения спазма сосудов. Распространяющийся ПД в гладкомышечных волокнах возникает только в том случае, если приложенный стимул возбуждает одновременно минимальное число мышечных волокон. Возбуждение распространяется посредством локальных токов. Потенциал действия может распространяться на соседние волокна через участки области контактов (нексусы) с относительно малым сопротивлением. Распространение ПД на определенное расстояние зависит от силы приложенного стимула. Скорость проведения ПД значительно меньше, чем в скелетных мышцах (10 см/сек), и тем больше, чем меньше число межклеточных переключений. Потенциал действия в гладкой мышце, как и в скелетной мускулатуре, имеет пусковое значение для начала сократительного процесса. Ионы Са++ здесь также инициируют процесс сокращения. Однако в большинстве гладких мышц саркоплазматический ретикулум развит плохо, количество кальция, запасаемого внутри клетки, невелико, поэтому значительно сильнее выражена зависимость сокращения от внеклеточной концентраций ионов Ca++, которые проникают внутрь мышечного волокна во время генерации потенциала действия. Ведущую роль в регуляции внутриклеточного кальция у большинства гладкомышечных клеток играет транспортные системы поверхностной мембраны: 1. Система подвижных переносчиков, обменивающих внутриклеточный Са++ на наружный Nа+; 2. Кальциевый насос (Са++ - АТФ-аза), использующий энергию АТФ для переноса ионов Са ++ в межклеточную среду. Латентный период одиночного сокращения гладкой мышцы больше, чем скелетной мышцы, и составляет 0, 25- 1, 00 сек, продолжительность сократительного ответа гладкой мышцы также велика (для мышц желудка кролика – 5 сек, лягушки - 1 мин и более). Особенно медленно протекает расслабление. Волна сокращения распространяется с той же скоростью, что и волна возбуждения (2 - 10 см/сек), но эта медлительность сократительной деятельности гладких мышц сочетается с большой их силой. Вследствие замедленного сокращения гладкая мышца даже при редких ритмических сокращениях (10 -12 раздражений в минуту) легко переходит в длительное состояние стойкого сокращения, напоминающее тетанус скелетных мышц. При этом энергетические расходы очень малы в отличие от тетануса поперечнополосатых мышц.
1.9. Нервно- мышечная передача возбуждения. Популярное:
|
Последнее изменение этой страницы: 2016-05-29; Просмотров: 2111; Нарушение авторского права страницы