|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Механизм проведения возбуждения по нервным и мышечным волокнам. Лабильность
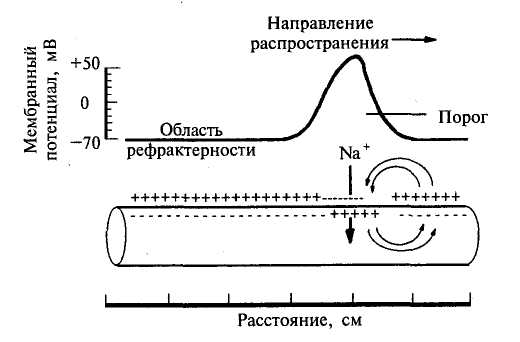
Механизм распространение возбуждения по нервным и мышечным волокнам осуществляется при помощи местных токов, возникающих между возбужденными и невозбужденными участками мембраны. Теория проведения возбуждения при помощи местных токов была сформулирована Германом в 1899 году и получила в настоящее время экспериментальное подтверждение. На рис.15 стрелками показано направление тока, возникающего между возбужденным (А) и соседним невозбужденным (Б) участками. Распространение местных токов по длине волокна определяется ее кабельными свойствами. Направление же местного тока таково, что он деполяризует соседний с активным (А) покоящийся (Б) участок мембраны. В итоге в участке Б уменьшается мембранный потенциал, т.е. происходит деполяризация, которая при достижении критического уровня вызывает возникновение ПД, и теперь уже возбужденный участок Б становится способным активировать соседний с ним покоящийся участок.
Рис. 15. Механизм распространения возбуждения с помощью местных токов после прохождения потенциала действия в нервном волокне возникает зона невозбудимости, или реф-рактерности.
В нервных мякотных волокнах, покрытых миелиновой оболочкой, ПД распространяется очень быстро, вследствие того, что возбуждение возникает только в перехватах Ранвье. К миелинизированным волокнам относятся чувствительные и двигательные волокна, а также часть волокон вегетативной нервной системы. На рис.16 показано строение миелинизированного нервного волокна. Миелиновая оболочка создается Шваннновской клеткой, многократно обертывающей осевой цилиндр волокна, поверхность которого образована плазматической мембраной. Свободные от миелиновой оболочки участки мембраны шириной, примерно, 1мкм называют перехватами Ранвье. В нервных волокнах, диаметром 10-20 мкм, длина межперехватных участков составляет 1-2 мм. Миелиновая оболочка участвует в регуляции обмена веществ и росте осевого цилиндра, выполняет функцию электрического изолятора, благодаря высокому сопротивлению и обеспечивает более экономное и быстрое проведение возбуждения.
А.
Рис. 4.1. Непрерывное распространение ПД в нервном волокне: 1 — возникновение ПД в нервном волокне и распространение ПД в обе стороны от места возникновения. В области деполяризации мембраны (1) преобладает входящий в клетку натриевый ток; 2 — соседняя область, в которой локальный ток от области ПД вызывает деполяризацию до критического уровня
Рис. 4.2. Сальтаторное распространение ПД в миелиновых нервных волокнах. Возникновение ПД в перехвате Ранвье среднего участка волокна и распространение ПД в обе стороны от места возникновения. Стрелками показаны токи на электротоническом этапе распространения ПД. В области перехватов Na+ движется в клетку, К+ — из клетки
Рис. 3.17. Схемы распространения ПД по нервным клеткам: а — общая схема; б — распространение ПД по неми-елинизированному (безмякотному) волокну (1, 2, 3, 4 — участки мембраны нейрона); в — распространение ПД по миелинизированному волокну
Рис. 16. Строение и распространение возбуждения по миелинизированному волокну. А. Образование миелинового чехла на аксоне. 1 — наматывание слоев миелина; 2 — аксон; 3 — олигодендроцит; 4 — перехват Ранвье; 5 — слой миелиновой оболочки. Б. Миелиновое нервное волокно. В. Распространение возбуждения
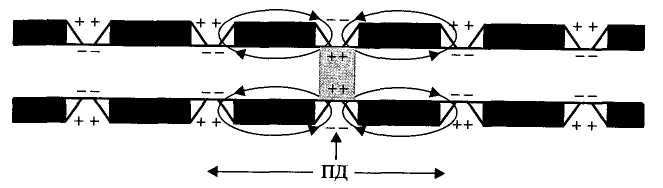
Участки между перехватами, покрытые миелином являются практически невозбудимыми. Перепрыгивание нервного импульса с одного перехвата на другой называют сальтаторное проведение возбуждения. Оно связано с высокой плотностью натриевых каналов в перехватах (до 1000 на 1 мкм мембраны). Сальтаторное проведение возбуждения от одного перехвата до другого оказывается возможным только потому, что амплитуда ПД в каждом перехвате в 5-6 раз превышает пороговую величину, необходимую для возбуждения соседнего перехвата Ранвье. Предположение о скачкообразном распространении возбуждения в нервных волокнах впервые было высказано Вериго в 1899 году. Возбуждение по мякотному нервному волокну проводится с большой скоростью без затухания (декремента), а интенсивность его одинакова в любом участке волокна. Расход же энергии при проведении возбуждения по миелинизированным волокнам невелик
Рис.17. Проведенеие возбуждения по миелинизированным нервным волокнам.
В мышечных и безмякотных волокнах возбуждение осуществляется непрерывно от точки к точке. Такое распространение возбуждения происходит с затуханием. Безмякотные волокна у позвоночных животных принадлежат в основном симпатической нервной системе. Они не имеют миелиновой оболочки, однако изолированы друг от друга Шванновскими клетками. Нервы обычно состоят из миелинизированных и немиелинизированных волокон в различном соотношении. В нервах иннервирующих скелетные мышцы преобладают миелинизированные волокна, а в нервах вегетативной нервной системы, например, в блуждающем нерве количество безмякотных волокон достигает 80-95%. Современная техника исследования позволяет точно определить скорость проведения возбуждения путем регистрации с помощью осциллографа латентного периода возникновения ПД при раздражении нерва с разных участков. Скорость проведения возбуждения зависит от диаметра волокна и структуры его мембраны - чем толще волокно, тем больше скорость проведения возбуждения в нем. В настоящее время нервные волокна по скорости проведения возбуждения, длительности различных фаз потенциала действия и строению принято подразделять на три основных типа, обозначаемых буквами А, Б и С.
Таблица 2. Свойства различных нервных волокон теплокровных
Рис. 30. Сокращения икроножной мышцы при разной частоте раздражения (по Е. Е. Жукову). Одиночное сокращение показано пунктиром. Цифры над кривыми — частоты стимулов. При частотах 20—35 в секунду — зубчатый тетанус, при частоте 1 15 в секунду — гладкий тетанус.
Таблица 3 Свойства различных нервных волокон теплокровных
Для сохранения функции нервного волокна необходима его связь с телом. При перерезке нервного волокна его периферический конец, отделенный от тела клетки перерождается и дегенерирует (у теплокровных животных через 2 - 3 дня), миелиновая оболочка скапливается в виде капель жира и рассасывается. При этом нарушается функция иннервируемого органа. Центральная же часть нервного волокна способна к регенерации. Регенерация нерва происходит очень медленно. Лабильность. Нервные волокна обладают разной лабильностью (функциональной подвижностью). Показателем лабильности является максимальное число потенциалов действия, которое способно возбудимое образование генерировать в 1с в соответствии с ритмом раздражения. Она определяется скоростями процессов изменений ионной проводимости, лежащих в основе абсолютной и относительной рефрактерности. При увеличении частоты ритмического раздражения лабильность ткани повышается, т.е. ткань отвечает более высокой частотой возбуждения по сравнению с исходной частотой. Это явление было открыто в 1923 году А.А. Ухтомским и получило название усвоение ритма раздражения. Лабильность нервных волокон колеблется от нескольких десятков до 500 импульсов в секунду, а в ряде случаев до 1000 и более. Наиболее высокой лабильностью обладают толстые миелинизированные волокна. Лабильность мышцы - около 200имп/сек, нервно-мышечного синапса- порядка 100 имп/сек. Существуют агенты, например, местные анестетики, способные замедлять реактивацию натриевых каналов и потому увеличивающие длительность фазы относительной рефрактерности. Так, если участок нервного волокна подвергнуть воздействию малых концентраций новокаина (или какого – либо другого местного анестетика), то волокно утрачивает способность проводить высокочастотные разряды импульсов, тогда как низкочастотные разряды еще продолжают проходить. Сходным образом влияет на процесс реактивации повышение концентрации ионов K+в окружающей нервные волокна жидкости. Обнаружены агенты, снижающие возбудимость (в результате блокады натриевых каналов), но не влияющие на длительность рефрактерной фазы. К числу таких агентов относится специфический блокатор натриевых каналов - тетродотоксин. Таким образом, различные по характеру своего воздействия агенты способны вызывать внешне сходные изменения возбудимости и проведения нервных импульсов. Обмен веществ в нервном волокне относительно невелик. О динамике обмена веществ нерва в покое и при возбуждении можно судить по его теплопродукции, которую впервые удалось зарегистрировать Хиллу в 1926 г. Деятельное состояние нерва характеризуется увеличением выделения тепла - повышением теплопродукции. Различают фазу начального теплообразования, длящуюся при одиночном раздражении в течение десятых долей секунды и фазу запаздывающего теплообразования, продолжающуюся в течение минуты. Исследования при помощи высокочувствительных приборов показали, что начальное теплообразование сопровождает процесс возбуждения (генерацию ПД), а запаздывающее теплообразование связано с восстановительными процессами. При возбуждении нервных волокон выделяется значительно меньше тепла по сравнению с количеством энергии, выделяемой мышцей. Деятельное состояние нерва характеризуется увеличением потребления аденозинтрифосфата (АТФ) и креатинфосфата (КФ), выделением молочной кислоты, усилением белкового обмена, обмена нуклеиновых кислот и фосфолипидов. Нервные волокна характеризуются относительной неутомляемостью, которая связана, прежде всего, с уровнем обмена веществ. Энергетические траты при возбуждении нервных волокон связаны главным образом с работой натрий - калиевого насоса.
Популярное:
|
Последнее изменение этой страницы: 2016-05-29; Просмотров: 2654; Нарушение авторского права страницы