|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Проводящие пути головного и спинного мозгаСтр 1 из 26Следующая ⇒
Системы нервных волокон, проводящих импульсы от кожи и слизистых оболочек, внутренних органов и органов движения к различным отделам спинного и головного мозга, в частности к коре полушарий большого мозга, называются восходящими, или чувствительными, афферентными, проводящими путями. Системы нервных волокон, передающих импульсы от коры или нижележащих ядер головного мозга через спинной мозг к рабочему органу (мышце, железе и др.), называются двигательными, или нисходящими, эфферентными, проводящими путями. Проводящие пути образованы цепями нейронов, причем чувствительные пути обычно состоят из трех нейронов, а двигательные - из двух. Первый нейрон всех чувствительных путей располагается всегда вне мозга, находясь в спинномозговых узлах или чувствительных узлах черепных нервов. Последний нейрон двигательных путей всегда представлен клетками передних рогов серого вещества спинного мозга или клетками двигательных ядер черепных нервов. Чувствительные пути. Спинной мозг проводит четыре вида чувствительности: тактильную (чувство прикосновения и давления), температурную, болевую и проприоцептивную (от рецепторов мышц и сухожилий, так называемое суставно-мышечное чувство, чувство положения и движения тела и конечностей). Основная масса восходящих путей проводит проприоцептивную чувствительность. Это говорит о важности контроля движений, так называемой обратной связи, для двигательной функции организма. Путь болевой и температурной чувствительности - латеральный спиноталамический путь. Первым нейроном этого пути являются клетки спинномозговых узлов. Периферические отростки их входят в состав спинномозговых нервов. Центральные отростки образуют задние корешки и идут в спинной мозг, оканчиваясь на клетках задних рогов (2-й нейрон). Отростки вторых нейронов через комиссуру спинного мозга переходят на противоположную сторону (образуют перекрест) и поднимаются в составе бокового канатика спинного мозга в продолговатый мозг. Там они примыкают к медиальной чувствительной петле и идут через продолговатый мозг, мост и ножки мозга к латеральному ядру таламуса, где переключаются на 3-й нейрон. Отростки клеток ядер таламуса образуют таламокортикальный пучок, проходящий через заднюю ножку внутренней капсулы к коре постцентральной извилины (область чувствительного анализатора). В результате того, что волокна по пути перекрещиваются, импульсы от левой половины туловища и конечностей передаются в правое полушарие, а от правой половины - в левое. Передний спиноталамический путь состоит из волокон, проводящих тактильную чувствительность, он проходит в переднем канатике спинного мозга. Пути мышечно-суставной (проприоцептивной) чувствительности направляются к коре полушарий большого мозга и в мозжечок, который участвует в координации движений. К мозжечку идут два спиномозжечковых пути - передний и задний. Задний спиномозжечковый путь (Флексига) начинается от клетки спинномозгового узла (1-й нейрон). Периферический отросток входит в состав спинномозгового нерва и заканчивается рецептором в мышце, капсуле суставов или связках. Центральный отросток в составе заднего корешка входит в спинной мозг и заканчивается в клетках ядра, расположенного у основания заднего рога (2-й нейрон). Отростки вторых нейронов поднимаются в дорсальной части бокового канатика этой же стороны и через нижние ножки мозжечка идут к клеткам коры червя мозжечка. Волокна переднего спиномозжечкового пути (Говерса) образуют перекрест дважды; в спинном мозге и в области верхнего паруса, а затем через верхние ножки мозжечка достигают клеток коры червя мозжечка. Проприоцептивный путь к коре больших полушарий представлен двумя пучками: нежным (тонким) и клиновидным. Нежный пучок (Голля) проводит импульсы от проприорецепторов нижних конечностей и нижней половины тела и лежит медиально в заднем канатике. Клиновидный пучок (Бурдаха) примыкает к нему снаружи и несет импульсы от верхней половины туловища и от верхних конечностей. Второй нейрон этого пути лежит в одноименных ядрах продолговатого мозга. Их отростки образуют перекрест в продолговатом мозге и соединяются в пучок, называемый медиальной чувствительной петлей. Она доходит до латерального ядра таламуса (3-й нейрон). Отростки третьих нейронов через внутреннюю капсулу направляются в чувствительную и частично двигательную зоны коры. Двигательные пути представлены двумя группами. 1. Пирамидные (кортикоспинальный и кортико-ядерный, или кортикобульбарный) пути, проводящие импульсы от коры к двигательным клеткам спинного и продолговатого мозга, являющиеся путями произвольных движений. 2. Экстрапирамидные, рефлекторные двигательные пути, входящие в состав экстрапирамидной системы. Пирамидный, или кортикоспинальный путь начинается от больших пирамидных клеток (Беца) коры верхних 2/3 предцентральной извилины и околоцентральной дольки, проходит через внутреннюю капсулу основание ножек мозга, основание моста, пирамиды продолговатого мозга. На границе со спинным мозгом он разделяется на боковой и передний пирамидные пучки. Боковой (большой) образует перекрест и спускается в боковом канатике спинного мозга, заканчиваясь на клетках переднего рога. Передний не перекрещивается и идет в переднем канатике. Образуя посегментный перекрест, его волокна также заканчиваются на клетках переднего рога. Отростки клеток переднею рога образуют передний корешок, двигательную порцию спинномозгового нерва и заканчиваются в мышце двигательным окончанием. Кортико-ядерный путь начинается в нижней трети предцентральной извилины, идет через колено (изгиб) внутренней капсулы и заканчивается на клетках двигательных ядер черепных нервов противоположной стороны. Отростки клеток двигательных ядер образуют двигательную порцию соответствующего нерва. К рефлекторным двигательным путям (экстрапирамидным) относятся красноядерно-спинномозговой (руброспинальный) путь - от клеток красного ядра среднего мозга, тектоспинальный путь - от ядер холмиков пластинки крыши среднего мозга (четверохолмия), связанный со слуховыми и зрительными восприятиями, и вестибулоспинальный - от вестибулярных ядер из ромбовидной ямки, связанный с поддержанием равновесия тела.
Раздел «Физиология» портала http: //medicinform.net Физиология спинного мозга Спинному мозгу присущи две функции: рефлекторная и проводниковая. Как рефлекторный центр спинной мозг способен осуществлять сложные двигательные и вегетативные рефлексы. Афферентными - чувствительными - путями он связан с рецепторами, а эфферентными - со скелетной мускулатурой и всеми внутренними органами. Длинными восходящими и нисходящими путями спинной мозг соединяет двусторонней связью периферию с головным мозгом. Афферентные импульсы по проводящим путям спинного мозга проводятся в головной мозг, неся ему информацию об изменениях во внешней и внутренней среде организма. По нисходящим путям импульсы от головного мозга передаются к эффекторным нейронам спинного мозга и вызывают или регулируют их деятельность.
Рефлекторная функция. Нервные центры спинного мозга являются сегментарными, или рабочими, центрами. Их нейроны непосредственно связаны с рецепторами и рабочими органами. Кроме спинного, мозга, такие центры имеются в продолговатом и среднем мозге. Надсегментарные центры, например промежуточного мозга, коры больших полушарий, непосредственной связи с периферией не имеют. Они управляют ею посредством сегментарных центров. Двигательные нейроны спинного мозга иннервируют все мышцы туловища, конечностей, шеи, а также дыхательные мышцы - диафрагму и межреберные мышцы. Помимо двигательных центров скелетной мускулатуры, в спинном мозге находится ряд симпатических и парасимпатических вегетативных центров. В боковых рогах грудного и верхних сегментах поясничного отделов спинного мозга расположены спинальные центры симпатической нервной системы, иннервирующие сердце, сосуды, потовые железы, пищеварительный тракт, скелетные мышцы, т.е. все органы и ткани организма. Именно здесь лежат нейроны, непосредственно связанные с периферическими симпатическими ганглиями. В верхнем грудном сегменте, находится симпатический центр расширения зрачка, в пяти верхних грудных сегментах - симпатические сердечные центры. В крестцовом отделе спинного мозга заложены парасимпатические центры, иннервирующие органы малого таза (рефлекторные центры мочеиспускания, дефекации, эрекции, эякуляции). Спинной мозг имеет сегментарное строение. Сегментом называют такой отрезок, который дает начало двум парам корешков. Если у лягушки перерезать на одной стороне задние корешки, а на другой передние, то, лапки на стороне, где перерезаны задние корешки, лишаются чувствительности, а на противоположной стороне, где перерезаны передние корешки, окажутся парализованными. Следовательно, задние корешки спинного мозга являются чувствительными, а передние - двигательными. В опытах с перерезкой отдельных корешков было установлено, что каждый сегмент спинного мозга иннервирует три поперечных отрезка, или метамера, тела: свой собственный, один выше и один ниже. Следовательно, каждый метамер тела получает чувствительные волокна от трех корешков и, для того чтобы лишить чувствительности участок тела, необходимо перерезать три корешка (фактор надежности). Скелетные мышцы также получают двигательную иннервацию от трех соседних сегментов спинного мозга. Каждый спинальный рефлекс имеет свое рецептивное поле и свою локализацию (место нахождения), свой уровень. Так, например, центр коленного рефлекса находится во II - IV поясничном сегменте; ахиллова - в V поясничном и I - II крестцовых сегментах; подошвенного - в I - II крестцовом, центр брюшных мышц - в VIII - XII грудных сегментах. Важнейшим жизненно важным центром спинного мозга является двигательный центр диафрагмы, расположенный в III - IV шейных сегментах. Повреждение его ведет к смерти вследствие остановки дыхания. Для изучения рефлекторной функции спинного мозга приготовляют спинальное животное - лягушку, кошку или собаку, делая поперечную перерезку спинного мозга ниже продолговатого. Спинальное животное в ответ на раздражение осуществляет оборонительную реакцию - сгибание или разгибание конечности, чесательный рефлекс - ритмическое сгибание конечностей, проприоцептивные рефлексы. Если спинальную собаку поднять за переднюю часть туловища и слегка надавить ей на подошву задней лапки, то возникает шагательный рефлекс - ритмическое, поочередное сгибание и разгибание лап. Проводниковая функция спинного мозга. Спинной мозг выполняет проводниковую функцию за счет восходящих и нисходящих путей, проходящих в белом веществе спинного мозга. Эти пути связывают отдельные сегменты спинного мозга друг с другом, а также с головным мозгом. Спинальный шок. Перерезка или травма спинного мозга вызывает явление, получившее название - спинального шока (шок в переводе с английского означает удар). Спинальный шок выражается в резком падении возбудимости и угнетении деятельности всех рефлекторных центров спинного мозга, расположенных ниже места перерезки. Во время спинального шока раздражители, обычно вызывающие рефлексы, оказываются недействительными. Укол лапы не вызывает сгибательного рефлекса. В то же время деятельность центров, расположенных выше перерезки, сохраняется. Обезьяна, которой перерезка спинного мозга была сделана в области верхних грудных сегментов, после того как пройдет наркоз, передними лапами берет банан, чистит его, подносит ко рту и съедает. После перерезки исчезают не только скелетно-моторные рефлексы, но и вегетативные. Снижается кровяное давление, отсутствуют сосудистые рефлексы, акты дефекации и микции (мочеиспускания). Продолжительность шока различна у животных, стоящих на различных ступенях эволюционной лестницы. У лягушки шок продолжается 3 - 5 мин, у собаки - 7 - 10 дней, у обезьяны - больше 1 месяца, у человека - 4 - 5 мес. Шок у человека нередко наблюдается как последствие бытовых или военных травм. Когда шок проходит, рефлексы восстанавливаются. Причиной спинального шока является выключение вышерасположенных отделов головного мозга, оказывающих на спинной мозг активирующее влияние, в котором большая роль принадлежит ретикулярной формации ствола мозга. Физиология продолговатого мозга Продолговатый мозг, так же как и спинной, выполняет две функции - рефлекторную и проводниковую. Из продолговатого мозга и моста выходят восемь пар черепных нервов (с V по XII) и он, так же как и спинной мозг, имеет прямую чувствительную и двигательную связь с периферией. По чувствительным волокнам он получает импульсы - информацию от рецепторов кожи головы, слизистых оболочек глаз, носа, рта (включая вкусовые рецепторы), от органа слуха, вестибулярного аппарата (органа равновесия), от рецепторов гортани, трахеи, легких, а также от интерорецепторов сердечно-сосудистой системы и системы пищеварения. Через продолговатый мозг осуществляются многие простые и сложнейшие рефлексы, охватывающие не отдельные метамеры тела, а системы органов, например системы пищеварения, дыхания, кровообращения. Рефлекторную деятельность продолговатого мозга можно наблюдать на бульбарной кошке, т. е. кошке, у которой произведена перерезка ствола мозга выше продолговатого. Рефлекторная деятельность такой кошки сложна и многообразна. Через продолговатый мозг осуществляются следующие рефлексы: Защитные рефлексы: кашель, чиханье, мигание, слезоотделение, рвота. Пищевые рефлексы: сосание, глотание, сокоотдение (секреция) пищеварительных желез. Сердечно-сосудистые рефлексы, регулирующие деятельность сердца и кровеносных сосудов. В продолговатом мозге находится автоматически работающий дыхательный центр, обеспечивающий вентиляцию легких. В продолговатом мозге расположены вестибулярные ядра. От вестибулярных ядер продолговатого мозга начинается нисходящий вестибулоспинальный тракт, участвующий в осуществлении установочных рефлексов позы, а именно в перераспределении тонуса мышц. Бульбарная кошка ни стоять, ни ходить не может, но продолговатый мозг и шейные сегменты спинного обеспечивают те сложные рефлексы, которые являются элементами стояния и ходьбы. Все рефлексы, связанные с функцией стояния, называются установочными рефлексами. Благодаря им животное вопреки силам земного притяжения удерживает позу своего тела, как правило, теменем кверху. Особое значение этого отдела центральной нервной системы определяется тем, что в продолговатом мозге находятся жизненно важные центры - дыхательный, сердечно-сосудистый, поэтому не только удаление, а даже повреждение продолговатого мозга заканчивается смертью. Помимо рефлекторной, продолговатый мозг выполняет проводниковую функцию. Через продолговатый мозг проходят проводящие пути, соединяющие двусторонней связью кору, промежуточный, средний мозг, мозжечок и спинной мозг. Физиология мозжечка Мозжечок не имеет прямой связи с рецепторами организма. Многочисленными путями он связан со всеми отделами центральной нервной системы. К нему направляются афферентные (чувствительные) проводящие пути, несущие импульсы от проприорецепторов мышц, сухожилий, связок, вестибулярных ядер продолговатого мозга, подкорковых ядер и коры больших полушарий. В свою очередь мозжечок посылает импульсы ко всем отделам центральной нервной системы. Функции мозжечка исследуют путем его раздражения, частичного или полного удаления и изучения биоэлектрических явлений. Последствия удаления мозжечка и выпадения его функции итальянский физиолог Лючиани охарактеризовал знаменитой триадой А - астазия, атония и астения. Последующие исследователи добавили еще один симптом - атаксия. Безмозжечковая собака стоит на широко расставленных лапах, совершая непрерывные качательные движения ( астазия ). У нее нарушено правильное распределение тонуса мышц сгибателей и разгибателей ( атония ). Движения плохо координированы размашисты, несоразмерны, резки. При ходьбе лапы забрасываются за среднюю линию ( атаксия ), чего не бывает у нормальных животных. Атаксия объясняется тем, что нарушается контроль движений. Выпадает и анализ сигналов от проприорецепторов мышц и сухожилий. Собака не может попасть мордой в миску с едой. Наклон головы вниз или в сторону вызывает сильное противоположное движение. Движения очень утомляют, животное, пройдя несколько шагов, ложится и отдыхает. Этот симптом называется астенией. С течением времени двигательные расстройства у безмозжечковой собаки сглаживаются. Она самостоятельно ест, походка ее почти нормальна. Только предвзятое наблюдение выявляет некоторые нарушения (фаза компенсации). Как показал Э.А. Асратян, компенсация функций происходит за счет коры головного мозга. Если у такой собаки удалить кору, то все нарушения выявляются снова и уже никогда не компенсируются. Мозжечок участвует в. регуляции движений, делая их плавными, точными, соразмерными. Как показали исследования Л.А. Орбели, у безмозжечковых собак нарушаются вегетативные функции. Константы крови, сосудистый тонус, работа пищеварительного тракта и другие вегетативные функции становится очень неустойчивыми, легко сдвигаются под влиянием тех или иных причин (приём пищи, мышечная работа, изменение температуры и др.). При удалении половины мозжечка нарушения двигательных функций наступают на стороне операции. Это объясняется тем, что проводящие пути мозжечка либо не перекрещиваются вовсе, либо перекрещиваются 2 раза. Физиология среднего мозга
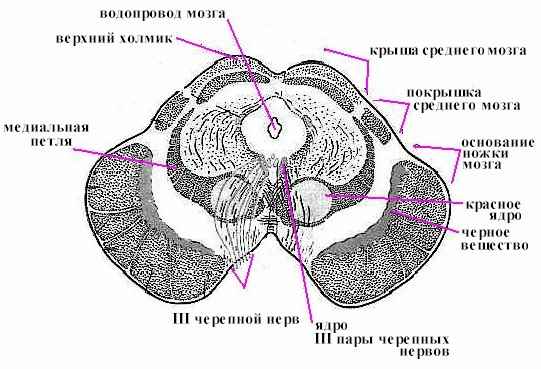
Средний мозг играет важную роль в регуляции мышечного тонуса и в осуществлении установочных и выпрямительных рефлексов, благодаря которым возможны стояние и ходьба. Роль среднего мозга в регуляции мышечного тонуса лучше всего наблюдать на кошке, у которой сделан поперечный разрез между продолговатым и средним мозгом. У такой кошки резко повышается тонус, мышц, особенно разгибателей. Голова запрокидывается назад, резко выпрямляются лапы. Мышцы настолько сильно сокращены, что попытка согнуть конечность заканчивается неудачей - она сейчас же распрямляется. Животное, поставленное на вытянутые, как палки, лапы, может стоять. Такое состояние называется децеребрационной ригидностью. Если разрез сделать выше среднего мозга, то децеребрационная ригидность не возникает. Примерно через 2 часа такая кошка делает усилие подняться. Сначала она поднимает голову, затем туловище, потом встает на лапы и может начать ходить. Следовательно, нервные аппараты регуляции мышечного тонуса и функции стояния и ходьбы находятся в среднем мозге. Явления децеребрационной ригидности объясняют тем, что перерезкой отделяются от продолговатого и спинного мозга красные ядра и ретикулярная формация. Красные ядра не имеют непосредственной связи с рецепторами и эффекторами, но они связаны со всеми отделами центральной нервной системы. К ним подходят нервные волокна от мозжечка, базальных ядер, коры полушарий большого мозга. От красных ядер начинается нисходящий руброспинальный тракт, по которому передаются импульсы к двигательным нейронам спинного мозга. Его называют экстрапирамидным трактом. Чувствительные ядра среднего мозга выполняют ряд важнейших рефлекторных функций. Ядра, находящиеся в верхних холмиках, являются первичными зрительными центрами. Они получают импульсы от сетчатки глаза и участвуют в ориентировочном рефлексе, т. е. повороте головы к свету. При этом происходит изменение ширины зрачка и кривизны хрусталика (аккомодация), способствующая ясному видению предмета. Ядра нижних холмиков являются первичными слуховыми центрами. Они участвуют в ориентировочном рефлексе на звук - поворот головы в сторону звука. Внезапные звуковые и световые раздражения вызывают сложную реакцию настораживания, мобилизующую животное на быструю ответную реакцию.
Физиология промежуточного мозга Главными образованиями промежуточного мозга являются таламус (зрительный бугор) и гипоталамус (подбугорная область). Таламус - чувствительное ядро подкорки. Его называют «коллектором чувствительности», так как к нему сходятся афферентные (чувствительные) пути от всех рецепторов, исключая обонятельные. Здесь находится третий нейрон афферентных путей, отростки которого заканчиваются в чувствительных зонах коры. Главной функцией таламуса является интеграция (объединение) всех видов чувствительности. Для анализа внешней среды недостаточно сигналов от отдельных рецепторов. Здесь происходит сопоставление информации, получаемой по различным каналам связи, и оценка ее биологического значения. В зрительном бугре насчитывается 40 пар ядер, которые подразделяются на специфические (на нейронах этих ядер заканчиваются восходящие афферентные пути), неспецифические (ядра ретикулярной формации) и ассоциативные. Через ассоциативные ядра таламус связан со всеми двигательными ядрами подкорки - полосатым телом, бледным шаром, гипоталамусом и с ядрами среднего и продолговатого мозга. Изучение функций зрительного бугра проводится путем перерезок, раздражения и разрушения. Кошка, у которой разрез сделан выше промежуточного мозга, резко отличается от кошки, у которой высшим отделом центральной нервной системы является средний мозг. Она не только поднимается и ходит, т. е. выполняет сложно координированные движения, но еще проявляет все признаки эмоциональных реакций. Легкое прикосновение вызывает злобную реакцию. Кошка бьет хвостом, скалит зубы, рычит, кусается, выпускает когти. У человека зрительный бугор играет существенную роль в эмоциональном поведении, характеризующемся своеобразной мимикой, жестами и сдвигами функций внутренних органов. При эмоциональных реакциях повышается давление, учащаются пульс, дыхание, расширяются зрачки. Мимическая реакция человека является врожденной. Если пощекотать нос плода 5 – 6 мес., то можно видеть типичную гримасу неудовольствия (П.К. Анохин). При раздражении зрительного бугра у животных возникают двигательные и болевые реакции - визг, ворчание. Эффект можно объяснить тем, что импульсы от зрительных бугров легко переходят на связанные с ними двигательные ядра подкорки. В клинике симптомами поражения зрительных бугров являются сильная головная боль, расстройства сна, нарушения чувствительности как в сторону повышения, так и понижения, нарушения движений, их точности, соразмерности, возникновение насильственных непроизвольных движений. Гипоталамус является высшим подкорковым центром вегетативной нервной системы. В этой области расположены центры, регулирующие все вегетативные функции, обеспечивающие постоянство внутренней среды организма, а также регулирующие жировой, белковый, углеводный и водно-солевой обмен. В деятельности вегетативной нервной системы гипоталамус играет такую же важную роль, какую играют красные ядра среднего мозга в регуляции скелетно-моторных функций соматической нервной системы. Самые ранние исследования функций гипоталамуса принадлежат К лоду Бернару. Он обнаружил, что укол в промежуточный мозг кролика вызывает повышение температуры тела почти на 3°С. Этот классический опыт, открывший локализацию центра терморегуляции в гипоталамусе, получил название теплового укола. После разрушения гипоталамуса животное становится пойкилотермным, т. е. теряет способность удерживать постоянство температуры тела. В холодной комнате температура тела понижается, а в жаркой повышается. Позднее было установлено, что почти все органы, иннервируемые вегетативной нервной системой, могут быть активированы раздражением подбугорной области. Иными словами, все эффекты, которые можно получить при раздражении симпатических и парасимпатических нервов, получаются при раздражении гипоталамуса. В настоящее время для раздражения различных структур мозга широко применяется метод вживления электродов. С помощью особой, так называемой стереотаксической техники, через трепанационное отверстие в черепе вводят электроды в любой заданный участок мозга. Электроды изолированы на всем протяжении, свободен только их кончик. Включая электроды в цепь, можно узко локально раздражать те или иные зоны. При раздражении передних отделов гипоталамуса возникают парасимпатические эффекты - усиление движений кишечника, отделение пищеварительных соков, замедление сокращений сердца и др. при раздражении задних отделов наблюдаются симпатические эффекты - учащение сердцебиения, сужение сосудов, повышение температуры тела и др. Следовательно, в передних отделах подбугорной области располагаются парасимпатические центры, а в задних - симпатические. Так как раздражение при помощи вживленных электродов производится на целом животном, без наркоза, то представляется возможность судить о поведении животного. В опытах Андерсена на козе с вживленными электродами был найден центр, раздражение которого вызывает неутолимую жажду, - центр жажды. При его раздражении коза могла выпивать до 10 л воды. Раздражением других участков можно было заставить сытое животное есть (центр голода). Широкую известность получили опыты испанского ученого Дельгадо на быке с электродом, вживленным в центр страха: Когда на арене разъяренный бык бросался на тореадора, включали раздражение, и бык отступал с ясно выраженными признаками страха. Американский исследователь Д. Олдз предложил модифицировать метод - предоставить возможность самому животному замыкать, что неприятных раздражений животное будет избегать и, наоборот, стремиться повторять приятные. Опыты показали, что имеются структуры, раздражение которых вызывает безудержное стремление к повторению. Крысы доводили себя до истощения, нажимая на рычаг до 14000 раз! Кроме того, обнаружены структуры, раздражение которых, по-видимому, вызывает крайне неприятное ощущение, так как крыса второй раз избегает нажать на рычаг и убегает от него. Первый центр, очевидно, является центром удовольствия, второй - центром неудовольствия. Чрезвычайно важным для понимания функций гипоталамуса явилось открытие в этом отделе мозга рецепторов, улавливающих изменения температуры крови (терморецепторы), осмотического давления (осморецепторы) и состава крови (глюкорецепторы). С рецепторов, обращенных в кровь, возникают рефлексы, направленные на поддержание постоянства внутренней среды организма - гомеостаза. «Голодная кровь», раздражая глюкорецепторы, возбуждает пищевой центр: возникают пищевые реакции, направленные на поиск и поедание пищи. Одним из частых проявлений заболевания гипоталамуса в клинике является нарушение водно-солевого обмена, проявляющееся в выделении большого количества мочи с низкой плотностью. Заболевание носит название несахарного мочеизнурения. Подбугорная область тесно связана с деятельностью гипофиза. В крупных нейронах надзрительного и околожелудочкового ядер гипоталамуса образуются гормоны - вазопрессин и окситоцин. По аксонам гормоны стекают к гипофизу, где накапливаются, а затем поступают в кровь. Иное взаимоотношение между гипоталамусом и передней долей гипофиза. Сосуды, окружающие ядра гипоталамуса, объединяются в систему вен, которые спускаются к передней доле гипофиза и здесь распадаются на капилляры. С кровью к гипофизу поступают вещества - релизинг-факторы, или освобождающие факторы, стимулирующие образование гормонов в передней его доле. Ретикулярная формация. В стволе мозга - продолговатом, среднем и промежуточном мозге, между его специфическими ядрами находятся скопления нейронов с многочисленными сильно ветвящимися отростками, образующими густую сеть. Эта система нейронов получила название сетчатого образования, или ретикулярной формации. Специальные исследования показали, что все так называемые специфические пути, проводящие определенные виды чувствительности от рецепторов к чувствительным зонам коры головного Мозга, дают в стволе мозга ответвления, заканчивающиеся на клетках ретикулярной формации. Потоки импульсов с периферии от экстеро-, интеро- и проприорецепторов. поддерживают постоянное тоническое возбуждение структур ретикулярной формации. От нейронов ретикулярной формации начинаются неспецифические пути. Они поднимаются вверх к коре головного мозга и подкорковым ядрам и спускаются вниз к нейронам спинного мозга. В чем же состоит функциональное значение этой своеобразной системы, не имеющей своей территории, располагающейся между специфическими соматическими и вегетативными ядрами ствола мозга? Методом раздражения отдельных структур ретикулярной формации удалось раскрыть ее функцию как регулятора функционального состояния спинного и головного мозга, а также важнейшего регулятора мышечного тонуса. Роль ретикулярной формации в деятельности центральной нервной системы сравнивают с ролью регулятора в телевизоре. Не давая изображения, он может менять громкость звука и освещенность. Раздражение ретикулярной формации, не вызывая двигательного эффекта, изменяет имеющуюся деятельность, тормозя ее или усиливая. Если у кошки короткими, ритмическими раздражениями чувствительного нерва вызывать защитный рефлекс - сгибание задней лапки, а затем на этом фоне присоединить раздражение ретикулярной формации, то в зависимости от зоны раздражения эффект будет различен: спинальные рефлексы либо резко усилятся, либо ослабятся и исчезнут, т. е. затормозятся. Торможение возникает при раздражении задних отделов ствола мозга, а усиление рефлексов - при раздражении передних отделов. Соответствующие зоны ретикулярной формации получили название тормозящей и активирующей зон. На кору головного мозга ретикулярная формация оказывает активирующее воздействие, поддерживая состояние бодрствования и концентрируя внимание. Если у спящей кошки с вживленными в промежуточный мозг электродами включать раздражение ретикулярной формации, то кошка просыпается, открывает глаза. На электроэнцефалограмме видно, что исчезают медленные волны, характерные для сна, и появляются быстрые волны, свойственные состоянию бодрствования. Ретикулярная формация оказывает на кору головного мозга восходящее, генерализованное (охватывающее всю кору) активирующее влияние. По выражению И.П. Павлова, «подкорка заряжает кору». В свою очередь кора больших полушарий регулирует активность сетчатого образования.
Популярное:
|
Последнее изменение этой страницы: 2016-07-12; Просмотров: 1472; Нарушение авторского права страницы