|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Теория системной динамической локализации высших психических функций
[…] Теория системной динамической локализации высших психических функций сформировалась в борьбе с двумя основными направлениями в решении проблемы «мозг и психика»: узким локализационизмом (или психоморфологическим направлением) и антилокализационизмом (или концепцией эквипотенциальности мозга). […] Узкий локализационизм исходит из представлений о психической функции как о неразложимой на компоненты единой психической «способности» (говорить, писать, читать, считать и т. д.), которая должна быть целиком соотнесена с определенными морфологическими структурами мозга. Сам мозг, и прежде всего кора больших полушарий, рассматривается данным направлением как совокупность различных «центров», каждый из которых целиком «заведует» определенной психической способностью, в связи с чем поражение какого-либо мозгового «центра» ведет к необратимому нарушению (или выпадению) соответствующей способности. Локализация психической функции («способности») понимается как непосредственное соотнесение психического и морфологического (или непосредственное «наложение» психического на морфологическое), в связи с чем это направление и получило название «психоморфологического». Следует отметить, что и в настоящее время психоморфологические представления отнюдь не изжиты, прежде всего в сознании некоторых врачей-клиницистов, отождествляющих локализацию того или иного симптома нарушений психической функции с локализацией этой функции. Наиболее яркими и последовательными представителями этого направления в разное время были неврологи, изучавшие последствия локальных поражений головного мозга (П. Брока, К. Вернике, Ф. Галль, В. Бродбент, К. Кляйст и многие другие). Френологическая карта Ф. Галля и локализационная карта К. Кляйста представляют собой логическое завершение идей узкого локализационизма о работе коры больших полушарий как совокупности различных «центров психических способностей». Другое направление — антилокализационизм — сходно с узким локализационизмом и по своему отношению к психическим функциям как к неразложимым психическим «способностям», и по своему пониманию локализации как непосредственного соотнесения психического и морфологического. Однако мозг, и прежде всего кора больших полушарий, трактуется данным направлением как однородное (эквипотенциальное) целое, равноценное и равнозначное по отношению к психическим функциям во всех своих отделах. Психические функции («способности») связаны равномерно со всем мозгом (и прежде всего с корой больших полушарий), и любое его поражение приводит к пропорциональному величине патологического очага нарушению всех психических функций одновременно (или к общему ухудшению сложных «символических функций», по К. Гольдштейну). Степень нарушения психической функции не зависит от локализации поражения, а определяется только массой пораженного мозга. Наиболее яркими представителями данного направления были некоторые физиологи (П. Флуранс, Ф. Гольц, К. Лешли и др.) и психологи-идеалисты (А. Бергсон и другие представители Вюрцбургской школы). Клинические наблюдения за больными с локальными поражениями мозга как будто бы подкрепляли фактами оба указанных направления: с одной стороны, поражение отдельных участков мозга (преимущественно коры больших полушарий) приводит к различным нарушениям психических процессов, с другой — при локальных поражениях мозга нередко наблюдаются факты высокой компенсации возникших нарушений, указывающие на возможность осуществления нарушенных функций и другими отделами мозга. Ни та, ни другая концепция не могли объяснить эти противоречия. […] Наконец, история науки знает и откровенное отрицание проблемы локализации высших психических функций человека, т. е. самой возможности связывать мозг и психику. Этой откровенно идеалистической позиции придерживались такие крупные физиологи, как Г. Гельмгольц, Ч. Шеррингтон, Э. Эдриан, Р. Гранит — естествоиспытатели в науке, но идеалисты по своему философскому мировоззрению. В конце XX века таких взглядов придерживался крупнейший физиолог Дж. Экклз, который известен своими работами по изучению синаптической передачи импульсов. В своих монографиях «Мозг и психический опыт», «Лицом к лицу с реальностью» Дж. Экклз выступает как субъективный идеалист, считающий первичным реальность собственного сознания, субъективный опыт, а весь остальной внешний мир — вторичным и рассматривающий человеческое сознание как «акт творений бога», а материалистическое понимание сознания — как «болезнь науки». Теория системной динамической локализации высших психических функций человека создавалась в борьбе с этими направлениями, с опорой на достижения отечественной психологии, с одной стороны, и материалистической физиологии — с другой. В отечественной нейропсихологии было пересмотрено понятие «функция». Высшие психические функции как психологические образования стали рассматриваться с позиций отечественной психологической науки как социальные по генезу (т.е. прижизненные, обусловленные социально-историческими факторами); опосредованные психологическими орудиями (главным образом речью); системные по строению; осознанные, динамичные по своей организации (т.е. различные по психологической структуре на разных этапах онтогенеза); произвольные по способу управления. Таким образом, представление о недифференцированных психических функциях как о далее неразложимых «способностях» было заменено современными представлениями о психических функциях как о «психологических системах», обладающих сложным психологическим строением и включающих много психологических компонентов (звеньев, фаз и т.д.). Нейропсихология рассматривает психические функции как образования, имеющие сложную рефлекторную основу, детерминированную внешними стимулами, или как сложные формы приспособительной деятельности организма, направленной на решение определенных психологических задач. В отечественной нейропсихологии пересмотрено и понятие «локализация». Локализация психических функций рассматривается как системный процесс. Это означает, что психическая функция (как и физиологическая, например дыхание) соотносится с мозгом как определенная многокомпонентная система, различные звенья которой связаны с работой разных мозговых структур. А.Р. Лурия считал, что высшие психические функции как сложные функциональные системы не могут быть локализованы в узких зонах мозговой коры или в изолированных клеточных группах, а должны опираться на сложные системы совместно работающих зон, располагающихся в различных, иногда далеко отстоящих друг от друга участках мозга, каждая из которых вносит свой вклад в осуществление психических процессов. Системная локализация высших психических функций предполагает их многоэтапную иерархическую многоуровневую мозговую организацию. Это неизбежно вытекает из сложного многокомпонентного состава функциональных систем, на которые опираются высшие психические функции. Локализация высших психических функций характеризуется также динамичностью, изменчивостью. Этот принцип следует из основных свойств функциональных систем, опосредующих высшие психические функции: пластичности, изменчивости, взаимозаменяемости входящих в их состав звеньев, — что подтверждается клиническими, физиологическими и анатомическими данными. Обобщая результаты многолетних клинических наблюдений, И.Н. Филимонов сформулировал положение о функциональной многозначности мозговых структур, согласно которому многие из них при определенных условиях могут включаться в выполнение новых функций. Существуют многочисленные физиологические доказательства справедливости идеи о динамичности, изменчивости мозговой организации функций. К ним относятся прежде всего экспериментальные исследования П. К. Анохина и его учеников, показавшие, что не только относительно сложные поведенческие акты (пищедобывательные, оборонительные и др.), но и сравнительно простые физиологические функции (например, дыхание) обеспечиваются сложными функциональными системами, где возможно замещение одних звеньев другими. Высшие психические функции человека обладают не только большей сложностью по сравнению с психическими функциями животных, а тем более с физиологическими функциями; они характеризуются качественными отличиями — осознанностью, опосредованностью: речью, произвольным способом управления, решающим значением социальных факторов в их формировании. Эти качественные отличия высших психических функций человека проявляются и в особенностях их мозговой организации. Еще Л.С. Выготский отмечал, что сравнительное изучение одних и тех же локальных мозговых поражений в детском и взрослом возрасте обнаруживает различные нарушения высших психических функций и что эти факты могут трактоваться лишь как следствие различий в мозговой организации высших психических функций у ребенка и взрослого. Формируясь прижизненно под влиянием социальных воздействий, высшие психические функции человека меняют свою психологическую структуру и, соответственно, свою мозговую организацию. Наиболее демонстративно это проявляется на примере речевых функций. Если у взрослого грамотного человека (правши) корковые поля средних отделов левого полушария играют ведущую роль в мозговом обеспечении речевых процессов, то у детей, еще не владеющих грамотой (до 5-6 лет), речевые процессы (понимание устной речи и активная речь) обеспечиваются мозговыми структурами и левого, и правого полушарий. Поражение корковых «речевых зон» левого полушария не ведет у них к выраженным речевым расстройствам. Мозг человека характеризуется четко выраженной межполушарной асимметрией, которую можно рассматривать в качестве важнейшей фундаментальной закономерности его работы. Межполушарная асимметрия проявляется и в моторных, и в сенсорных функциях, но наиболее отчетливо — в высших психических функциях. Межполушарные различия в мозговой организации высших психических функций многократно описаны в клинической и нейропсихологической литературе как различия симптомов и синдромов, связанных с поражением симметричных отделов больших полушарий. С позиций теории системной динамической локализации функций эти различия можно обозначить как принцип различной локализации (или мозговой организации) всех высших психических функций в левом и правом полушариях мозга или принцип латеральной специализации мозговой организации психических функций. Изучение конкретной мозговой организации каждой из высших психических функций (у детей и взрослых) в значительной степени является делом будущего, так же как и изучение качественных различий мозговой организации психических процессов, свойственных левому и правому полушариям. Однако современный уровень знаний о межполушарной асимметрии мозга, достигнутый нейропсихологией и другими науками (физиологией, анатомией, неврологией и др.), подтверждает бесспорное значение данного принципа для понимания общих закономерностей локализации высших психических функций у человека. Человеческий мозг характеризуется мощным развитием лобных долей. По площади, занимаемой корой лобных долей, человек превосходит всех остальных представителей животного мира. «Лобный мозг» у человека продолжает развиваться в онтогенезе и окончательно формируется лишь к 12-14 годам или даже позже. Являясь прежде всего мозговым субстратом произвольных, сложнопрограммируемых форм поведения, «лобный мозг» постепенно, по мере созревания включается в реализацию всех высших психических функций человека как одно из важнейших звеньев функциональных систем. С позиций теории системной динамической локализации высших психических функций человека эту закономерность можно сформулировать как принцип обязательного участия лобных префронтальных отделов коры больших полушарий в мозговом обеспечении высших психических функций. Этот принцип специфичен для мозговой организации всех сознательных форм психической деятельности человека. Итак, согласно теории системной динамической локализации высших психических функций человека каждая высшая психическая функция обеспечивается мозгом как целым, однако это целое состоит из высоко дифференцированных структур (систем, зон), каждая из которых вносит свой вклад в реализацию функции. Непосредственно с мозговыми структурами (факторами) следует связывать не всю психическую функцию и даже не отдельные ее звенья, а те физиологические процессы, которые осуществляются в этих мозговых структурах и обеспечивают реализацию определенных аспектов (параметров) функции. Нарушение этих физиологических процессов ведет к появлению первичных дефектов, а также взаимосвязанных с ними вторичных дефектов (первичных и вторичных нейропсихологических симптомов), составляющих в целом закономерное сочетание нарушений высших психических функций — определенный нейропсихологический синдром. Основные принципы строения мозга Мозг как субстрат психических процессов представляет собой единую суперсистему, единое целое, состоящее, однако, из дифференцированных отделов (участков или зон), которые выполняют различную роль в реализации психических функций. Это главное положение теории локализации высших психических функций человека опирается не только на сравнительно-анатомические, физиологические данные и результаты клинических наблюдений, но и на современные сведения об основных принципах строения мозга человека. Что такое мозг как субстрат высших психических функций? Какие отделы мозга играют ведущую роль в их реализации? Все данные (и анатомические, и физиологические, и клинические) свидетельствуют о ведущей роли коры больших полушарий в мозговой организации психических процессов. Кора больших полушарий (и прежде всего, новая кора) является наиболее дифференцированным по строению и функциям отделом головного мозга. В недавнем прошлом коре больших полушарий придавалось исключительное значение, ее считали единственным субстратом психических процессов. Подкорковым структурам отводилась вспомогательная роль, за ними признавались прежде всего энергетические, активационные функции. Однако по мере накопления знаний о подкорковых образованиях представления об их участии в реализации различных психических процессов изменились. В настоящее время общепризнанной стала точка зрения о важной и специфической роли не только корковых, но и подкорковых структур в психической деятельности при ведущем участии коры больших полушарий. Таким образом, все высшие психические функции имеют и горизонтальную (корковую), и вертикальную (подкорковую) мозговую организацию. Как известно, головной мозг (encephalon) — высший орган нервной системы — как анатомо-функциональное образование может быть условно подразделен на несколько уровней, каждый из которых осуществляет собственные функции. I уровень — кора головного мозга — осуществляет высшее управление чувствительными и двигательными функциями, преимущественное управление сложными когнитивными процессами. II уровень — базальные ядра полушарий большого мозга — осуществляет управление непроизвольными движениями и регуляцию мышечного тонуса. III уровень — гиппокамп, гипофиз, гипоталамус, поясная извилина, миндалевидное ядро — осуществляет преимущественное управление эмоциональными реакциями и состояниями, а также эндокринную регуляцию. IV уровень (низший) — ретикулярная формация и другие структуры ствола мозга — осуществляет управление вегетативными процессами (Р. Д. Синельников, Я. Р. Синельников, 1996). Головной мозг подразделяется на ствол, мозжечок и большой мозг. Как анатомическое образование большой мозг (cerebrum) состоит из двух полушарий — правого и левого (hemisphererum cerebri dextrum et sinistrum); в каждом из них объединяются три филогенетически и функционально различные системы: 1 ) обонятельный мозг (rhinencephalon); 2) базальные ядра (nuclii basales); 3) кора большого мозга (cortex cerebri) — конвекситальная, базальная, медиальная. В каждом полушарии имеется пять долей: 1) лобная (lobus frontalis); 2) теменная (lobus parietalis); 3) затылочная (lobus occipitalis); 4) височная (lobus temporalis); 5) островковая, островок (lobus insularis, insule). Как известно, у человека по сравнению с другими представителями животного мира существенно больше развиты филогенетически новые отделы мозга, и прежде всего кора больших полушарий. Кора большого мозга (cortex cerebri) — наиболее высокодифференцированный раздел нервной системы — подразделяется на следующие структурные элементы: ♦ древнюю (paleocortex); ♦ старую (archeocortex); ♦ среднюю, или промежуточную (mesocortex); ♦ новую (neocortex). У человека новая кора — наиболее сложная по строению — по протяженности составляет 96 % от всей поверхности полушарий. Наиболее типична для человека новая шестислойная кора, однако в разных отделах мозга число слоев различно. По морфологическим критериям выделены разные цитоархитектонические поля, характеризующиеся различным строением клеток. Наибольшее признание получила цитоархитектоническая карта полей Бродмана, согласно которой выделяется 52 поля. В пределах многих полей выделены подполя. В пределах новой коры у человека наибольшее развитие получили ассоциативные отделы. Одновременно отмечаются усложнение и дифференцировка ассоциативных таламических ядер, подкорковых узлов, а также филогенетически новых отделов мозгового ствола. Существенно более развиты у человека по сравнению со всеми представителями животного мира, включая и высших приматов, лобные доли мозга — как их корковые отделы, так и подкорковые связи. Ассоциативные отделы коры больших полушарий у человека не только больше по занимаемой площади, чем проекционные (в абсолютных и относительных размерах), но и характеризуются более тонким архитектоническим и нейронным строением. На основании анализа новых экспериментальных данных, полученных в Институте Мозга РАМН и в других научных учреждениях, а также обобщения огромного литературного материала О. С. Адриановым (1983 и др.) была разработана концепция структурно-системной организации мозга как субстрата психической деятельности. В соответствии с этой концепцией деятельность мозга обеспечивается проекционными, ассоциативными, интегративно-пусковыми и лимбико-ретикулярными системами, каждая из которых выполняет свои функции. Проекционные системы обеспечивают анализ и переработку соответствующей по модальности информации. Ассоциативные системы связаны с анализом и синтезом разномодальных возбуждений.
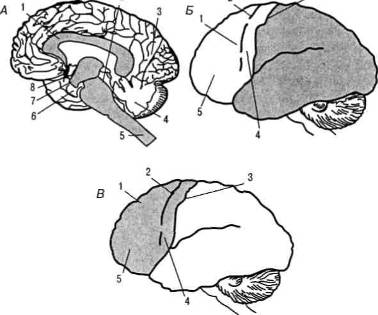
Рисунок: карта цитоархитектонических полей коры головного мозга: А — конвекситальная кора; Б — медиальная кора. Цифрами обозначены отдельные корковые поля; цифрами и буквами — подполя.
Для интегративно-пусковых систем характерен синтез возбуждений различной модальности с биологически значимыми сигналами и мотивационными влияниями, а также окончательная трансформация афферентных влияний в качественно новую форму деятельности, направленную на быстрейший выход возбуждений на периферию (т.е. на аппараты, реализующие конечную стадию приспособительного поведения). Лимбико-ретикулярные системы обеспечивают энергетические, мотивационные и эмоционально- вегетативные влияния. Все перечисленные выше системы мозга работают в тесном взаимодействии друг с другом по принципу либо одновременно, либо последовательно возбужденных структур. Работа каждой системы, а также процессы взаимодействия систем имеют не жестко закрепленный, а динамический характер. Эта динамика определяется особенностями поступающих афферентных импульсов и спецификой реакции организма. Установлено, что головной мозг человека обладает значительной изменчивостью. Различают этническую, половую, возрастную и индивидуальную изменчивость. Этнические различия, сохраняющиеся от поколения к поколению, относятся к общему весу (массе) головного мозга, его размерам, организации борозд и извилин. Считается, однако, что средний вес мозга, свойственный одной этнической группе, — весьма условный показатель, так как индивидуальная изменчивость может перекрывать средние величины. Масса мозга коррелирует с весом тела и формой черепа. Установлены различия между мужским и женским мозгом: 1375 г для мужчин и 1245 г для женщин — средние показатели веса мозга европейца. С возрастом масса мозга и морфологическое строение отдельных структур и проводящих волокон (мозолистого тела, передних комиссур и др.) изменяются, причем у женщин эти изменения менее заметны, чем у мужчин. С момента рождения головной мозг постепенно увеличивается и достигает максимальной массы к 20 годам; после 50 лет происходит постепенное уменьшение массы мозга (примерно на 30 г каждые 10 лет жизни). Описана значительная индивидуальная морфологическая изменчивость мозга. Это относится и к массе мозга, и к другим его характеристикам. Современная нейроанатомия признает существование пороговых значений веса мозга: по одним данным, минимальная масса мозга равна 900 г; по другим — 750-800 г (С.В. Савельев, 1996). При объеме мозга 246-622 см (микроцефалия) наблюдается явное снижение умственных способностей. Максимальная масса мозга здорового человека равна 2200-2300 г. Еще большая масса, как правило, является следствием патологического процесса (гидроцефалии и др.). Помимо веса индивидуальные морфологические различия относятся и к организации мозга. Существует высокая изменчивость в строении поверхности полушарий переднего мозга, что отражается в изменчивости строения его борозд и извилин. Достаточно велика индивидуальная изменчивость и подкорковых образований, что не связано ни с объемом мозга, ни с полом, ни с национальной принадлежностью. Так, объем подкорковых ядер (скорлупа, хвостатое ядро и др.) у разных людей может различаться в 2-3 раза. Весьма важным принципом структурной организации мозга как субстрата психической деятельности является также принцип иерархической соподчиненности различных систем мозга, соответственно которому уменьшается число степеней свободы каждой нижележащей системы и осуществляется управление одного уровня иерархии другими, а также контроль за этим управлением на основе прямых и обратных связей. Вместе с тем подобная иерархия допускает определенную избыточность в структурной организации мозга за счет вовлечения в ту или иную его функцию большого числа нервных элементов, что приводит к повышению надежности работы мозга и служит основой для компенсации функций при его поражениях. Принцип иерархии систем, как и другие принципы организации мозга, обеспечивает его интегративную целостную деятельность. Наконец, современная нейропсихология выдвигает как один из важнейших принципов структурно- системной организации мозга принцип многоуровневого взаимодействия вертикально организованных (подкорково-корковых) и горизонтально организованных (корково-корковых) путей проведения возбуждения, что создает широкие возможности для различных типов переработки (трансформации) афферентных сигналов и является одним из механизмов интегративной работы мозга. Данная концепция дает анатомическое обоснование двум основным принципам теории локализации высших психических функций, разработанной в нейропсихологии: ♦ принципу системной локализации функций (каждая психическая функция опирается на сложные взаимосвязанные структурно-функциональные системы мозга); ♦ принципу динамической локализации функции (каждая психическая функция имеет динамическую, изменчивую мозговую организацию, различную у разных людей и в разные возрастные периоды). […] В нейропсихологии на основе анализа клинических данных (т. е. изучения нарушений психических процессов при различных локальных поражениях мозга) была разработана общую структурно-функциональную модель работы мозга как субстрата психической деятельности. Эта модель, предложенная А.Р. Лурия, характеризует наиболее общие закономерности работы мозга как единого целого и является основой для объяснения его интегративной деятельности. Согласно данной модели, весь мозг может быть подразделен на три основных структурно-функциональных блока: I — энергетический блок, или блок регуляции уровня активности мозга; II — блок приема, переработки и хранения экстероцептивной (т. е. исходящей извне) информации; III — блок программирования, регуляции и контроля за протеканием психической деятельности. Каждая высшая психическая функция (или сложная форма сознательной психической деятельности) осуществляется при участии всех трех блоков мозга, вносящих свой вклад в ее реализацию. Они характеризуются определенными особенностями строения, физиологическими принципами, лежащими в основе их работы, и той ролью, которую они играют в осуществлении психических функций. Энергетический блок включает неспецифические структуры разных уровней: ♦ ретикулярную формацию ствола мозга; ♦ неспецифические структуры среднего мозга, его диэнцефальных отделов; ♦ лимбическую систему; ♦ медиобазальные отделы коры лобных и височных долей мозга.
Рисунок: структурно-функциональная модель интегративной работы мозга (по А.Р. Лурия, 1970): А) I блок — регуляции общей и избирательной неспецифической активации мозга, — включающий ретикулярные структуры ствола, среднего мозга и диэнцефальных отделов, а также лимбическую систему и медиобазальные отделы коры лобных и височных долей мозга: 1 — мозолистое тело, 2 — средний мозг, 3 — теменно-затылочная борозда, 4 — мозжечок, 5 — ретикулярная формация ствола, 6 — крючок, 7 — гипоталамус, 8 — таламус; Б) II блок — приема, переработки и хранения экстероцептивной информации, — включающий основные анализаторные системы (зрительную, кожно-кинестетическую, слуховую), корковые зоны которых расположены в задних отделах больших полушарий: 1 — премоторная область, 2 — прецентральная извилина, 3 — центральная извилина, 4 — моторная область, 5 — префронтальная область; В) III блок — программирования, регуляции и контроля за протеканием психической деятельности, — включающий моторные, премоторные и префронтальные отделы мозга с их двухсторонними связями. Обозначения те же, что и на рис. Б
Данный блок мозга регулирует два типа процессов активации: ♦ общие генерализованные изменения активации, являющиеся основой различных функциональных состояний; ♦ локальные избирательные изменения активации, необходимые для осуществления высших психических функций. Первый тип процессов активации связан с длительными тоническими сдвигами в активационном режиме работы мозга, с изменением уровня бодрствования. Второй тип процессов активации — это преимущественно кратковременные фазические изменения в работе отдельных структур (систем) мозга. Разные уровни неспецифической системы вносят свой вклад в обеспечение длительных тонических и кратковременных фазических процессов активации: ♦ нижние уровни неспецифической системы (ретикулярные отделы ствола и среднего мозга) обеспечивают преимущественно первый генерализованный тип процессов активации; ♦ расположенные выше уровни неспецифической системы (диэнцефальный, лимбический и особенно корковый) связаны преимущественно с регуляцией кратковременных фазических, избирательных форм процессов активации; ♦ медиобазальные отделы коры лобных долей больших полушарий обеспечивают регуляцию избирательных селективных форм процессов активации, которая осуществляется с помощью речевой системы. Первый тип процессов активации связан преимущественно с работой медленно действующей системы регуляции активности. Второй тип процессов активации обеспечивается механизмами быстродействующей активационной системы, регулирующей протекание различных ориентировочных реакций. Неспецифические структуры первого блока по принципу своего действия подразделяются на следующие типы: ♦ восходящие (проводящие возбуждение от периферии к центру); ♦ нисходящие (проводящие возбуждение от центра к периферии). Восходящие и нисходящие отделы неспецифической системы включают и активационные, и тормозные пути. В настоящее время установлено, что активационные и тормозные неспецифические механизмы являются достаточно автономными и независимыми по своей организации на всех уровнях, включая и кору больших полушарий. Анатомические особенности неспецифической системы состоят прежде всего в наличии в ней особых клеток, составляющих ретикулярную (сетчатую) формацию и обладающих, как правило, короткими аксонами, что объясняет сравнительно медленную скорость распространения возбуждения в этой системе. Однако в неспецифических структурах обнаружены и длинноаксонные клетки, участвующие в механизме быстрых активационных процессов. Корковые структуры первого блока (поясная кора, кора медиальных и базальных, или орбитальных, отделов лобных долей мозга) принадлежат по своему строению главным образом к коре древнего типа, состоящей из пяти слоев. Функциональное значение первого блока в обеспечении психических функций состоит, как уже говорилось выше, прежде всего в регуляции процессов активации, в обеспечении общего активационного фона, на котором осуществляются все психические функции, в поддержании общего тонуса ЦНС, необходимого для любой психической деятельности. Этот аспект работы блока имеет непосредственное отношение к процессам внимания — общего, неизбирательного и селективного, — а также сознания в целом. Внимание и сознание с энергетической точки зрения связаны с определенными уровнями активации. С качественной, содержательной точки зрения они характеризуются набором различных действующих систем и механизмов, обеспечивающих отражение разных аспектов внешнего и внутреннего мира. Помимо общих неспецифических активационных функций, первый блок мозга непосредственно связан с процессами памяти (в их модально-неспецифической форме), с запечатлением, хранением и переработкой разномодальной информации. Решающее значение этого блока в мнестической деятельности подтверждено многочисленными наблюдениями за больными с поражением срединных неспецифических структур мозга, причем высшие уровни этих структур связаны преимущественно с произвольными формами мнестической деятельности. Первый блок мозга является непосредственным мозговым субстратом различных мотивационных и эмоциональных процессов и состояний (наряду с другими мозговыми образованиями). Лимбические структуры мозга, входящие в этот блок (область гиппокампа, поясной извилины, миндалевидного ядра и др.), имеющие тесные связи с орбитальной и медиальной корой лобных и височных долей мозга, являются полифункциональными образованиями. Они участвуют в регуляции различных эмоциональных состояний, прежде всего сравнительно элементарных (базальных) эмоций (страха, удовольствия, гнева и др.), а также мотивационных процессов, связанных с различными потребностями организма. В сложной мозговой организации эмоциональных и мотивационных состояний и процессов лимбические отделы мозга занимают одно из центральных мест. Этот блок мозга воспринимает и перерабатывает разную интероцептивную информацию о состояниях внутренней среды организма и регулирует эти состояния с помощью нейрогуморальных, биохимических механизмов. Таким образом, первый блок мозга участвует в осуществлении любой психической деятельности, особенно в процессах внимания, памяти, регуляции эмоциональных состояний и сознания в целом. Второй блок — блок приема, переработки и хранения экстероцептивной (т. е. исходящей из внешней среды) информации — включает основные анализаторские системы: зрительную, слуховую и кожно-кинестическую, корковые зоны которых расположены в задних отделах больших полушарий головного мозга. Работа этого блока обеспечивает модально-специфические процессы, а также сложные интегративные формы переработки экстероцептивной информации, необходимой для осуществления высших психических функций. Модально-специфические (или лемнисковые) пути проведения возбуждения имеют иную, чем неспецифические пути, нейронную организацию, им присуща четкая избирательность, проявляющаяся в реагировании лишь на определенный тип раздражителей. Все основные анализаторные системы организованы по общему принципу: они состоят из периферического (рецепторного) и центрального отделов. Периферические отделы анализаторов осуществляют анализ и дискриминацию стимулов по их физическим качествам (интенсивности, частоте, длительности и т. п.). Центральные отделы анализаторов включают несколько уровней, последний из которых — кора больших полушарий. Центральные отделы анализируют и синтезируют стимулы не только по физическим параметрам, но и по сигнальному значению. В целом анализаторы — это аппараты, подготавливающие ответы организма на внешние раздражители. На каждом из уровней анализаторной системы происходит последовательное усложнение процессов переработки информации. Максимальной сложности и дробности процессы анализа и переработки информации достигают в коре больших полушарий. Анализаторные системы характеризуются иерархическим принципом строения, при этом нейронная организация их уровней различна. Кора задних отделов больших полушарий обладает рядом общих черт, позволяющих объединить ее в единый блок мозга. В ней выделяют «ядерные зоны» анализаторов и «периферию» (по терминологии И.П. Павлова), или первичные, вторичные и третичные поля (по терминологии А.В. Кэмпбелла). К ядерным зонам анализаторов относят первичные и вторичные поля, к периферии — третичные поля. В ядерную зону зрительного анализатора входят 17, 18 и 19-е поля, в ядерную зону кожно-кинестетического анализатора — 1, 2, 3-е, частично 5-е поля, в ядерную зону звукового анализатора — 41, 42 и 22-е поля, из них первичными полями являются 3, 17 и 41-е, остальные — вторичные. Первичные поля коры по своей цитоархитектонике принадлежат к коникортикальному, или пылевидному, типу, который характеризуется широким IV слоем с многочисленными мелкими зерновидными клетками. Эти клетки принимают и передают пирамидным нейронам III и V слоев импульсы, приходящие по афферентным проекционным волокнам из подкорковых отделов анализаторов. Так, первичное 17-е поле коры содержит в IV слое крупные звездчатые клетки, откуда импульсы переключаются на пирамидные клетки V слоя (клетки Кахала и клетки Майнерта). От пирамидных клеток первичных полей берут начало нисходящие проекционные волокна, поступающие в соответствующие двигательные центры местных двигательных рефлексов (например, глазодвигательных). Все первичные корковые поля характеризуются топическим принципом организации («точка в точку»), согласно которому каждому участку рецепторной поверхности (сетчатки, кожи, кортиевого органа) соответствует определенный участок в первичной коре, что и дало основание называть ее проекционной. Величина зоны представительства того или иного рецепторного участка в первичной коре зависит от функциональной значимости этого участка. Так, область fovea представлена в 17-м поле коры значительно более широко, чем другие области сетчатки. Популярное:
|
Последнее изменение этой страницы: 2016-07-12; Просмотров: 1111; Нарушение авторского права страницы