|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ИСТОРИЯ ИССЛЕДОВАНИЯ ПЛАЗМИДСтр 1 из 6Следующая ⇒
ВВЕДЕНИЕ
Плазмиды – внехромосомные генетические элементы, способные к автономному поддержанию в цитоплазме бактерий или существованию в интегрированном в хромосому состоянии, откуда они могут свободно выходить в цитоплазму (иногда с фрагментами хромосомы). Некоторые хромосомы могут распространяться в бактериальной популяции между ее членами. Плазмиды определяют ряд важных свойств бактерий: · Являются факторами фертильности – определяют донорский фенотип клетки. · Контролируют резистентность к антибиотикам, сульфаниламидам, катионам тяжелых металлов, бактериоцинам, бактериофагам, к сыворотке крови. · Чувствительность к бактериоцинам · Синтез тиамина, пролина, внеклеточной ДНКазы и др. · Синтез антибиотиков и бактериоцинов. · Метаболизм углеводов, углеводсодержащих соединений, галогеновых соединений, белков. · Фиксацию азота. · Продукцию токсинов, гемолизина, антигенов колонизации, капсулы. В последнее время природа факторов внехромосомной наследственности микроорганизмов приобрела особый интерес в связи с появлением данных о возможности использования плазмид в качестве векторов эукариотных генов. Такая возможность открывает неограниченные перспективы для генетического моделирования не только при решении проблем молекулярной биологии, но и в практическом аспекте, в частности в медицинской микробиологии и иммунологии (создание новых бактерийных профилактических и лечебных препаратов) и микробиологической промышленности. [3, c. 98] Большой опыт экспериментального мутагенеза на модели бактерий и вирусов способствовал раскрытию генетических и молекулярных механизмов регуляции функций внехромосомных элементов. Их способность включаться в хромосому и формировать комплексы «замещенных» плазмид широко используется в экспериментальной биологии и генетике. Замещенные плазмиды несут фрагменты хромосомы бактерии-хозяина и в автономном состоянии функционируют под контролем регуляторных механизмов бактериальной клетки. Расширение методических и технических возможностей экспериментальных исследований в области молекулярной биологии позволяет целенаправленно использовать генетические модели в решении важных практических задач. Определились реальные пути более гибкого вмешательства в процессы физиологически нормального генетического обмена у бактерий, осуществляемого с участием внехромосомных элементов, способствующих конъюгации, формированию рекомбинантов, передаче генетического материала путем трансдукции умеренными фагами, мобилизации нетрансмиссивных элементов плазмидами, имеющими в своей структуре «гены трансмиссивности», и сочетания с этими генами фрагментов хромосомы с последующим переносом вновь формирующихся структур и их ассоциаций в клетки реципиентов. Актуальное значение приобретает исследование механизмов взаимодействия внехромосомных элементов с хромосомой и между собой в естественных или сконструированных искусственно полиплазмидных системах. Подчинение этих систем общим регуляторным механизмам на уровне клетки и популяции микроорганизмов выдвигает новые проблемы: изучение специфических особенностей полиплазмидных популяций при наличии дополнительных генетических факторов, не обязательных для воспроизведения жизнеспособного потомства, и возможностей практического использования искусственно обогащенного генома популяций бактерий. В последнее десятилетие интенсивно накапливаются данные о генетической природе и биологических особенностях плазмид, с которыми непосредственно связана патологическая активность бактерий. Это – элементы Hly, Ent, Vir, сведения о которых в мало обобщены. Практическое значение в инфекционной патологии приобретают «вторичные» процессы при ожоговых заболеваниях и постхирургических осложнениях, возникающих в связи с неограниченно возрастающей множественностнной лекарственной устойчивостью возбудителей этих процессов, контролируемой трансмиссивными и нетрансмиссивными факторами инфекционной резистентности. Менее полно изучены, но не менее важны плазмиды, контролирующие патогенные свойства стафилококков, стрептококков, псевдомонад. [7, c. 163] В настоящее время на основе использования трансмиссивных эписом интенсивно разрабатывается новое направление исследований - «генетическая инженерия» и как специальный раздел этого направления - «генная инженерия». Последняя представляет собой область прикладной молекулярной генетики и биологии, развитие которой только начинается. Однако первоисточником «сырья» для осуществления конкретных задач конструирования новых биологически активных молекул являются внехромосомные элементы, способные функционировать в виде самостоятельных оперонов и репликонов. Они сохраняют эту функцию в гетерологичных системах микроорганизмов и, что особенно привлекает «биоинженеров», - в системах эукариотов. Целью данной работы является изучить роль плазмид в жизнедеятельности бактериальной клетки. В работе поставлены следующие задачи: 1. изучить историю исследования плазмид; 2. рассмотреть особенности идентификации и классификации плазмид; 3. рассмотреть генетическую и молекулярную организацию плазмид; 4. изучить особенности поддержания плазмид в бактериальных клетках; 5. выявить сущность конъюгационного переноса; 6. рассмотреть свойства бактерий, контролируемые бактериями. Исходя из поставленных задач, была определена структура работы. Данная курсовая работа включает введение, основную часть, состоящую из семи глав, заключение и список использованной литературы.
ИДЕНТИФИКАЦИЯ ПЛАЗМИД Идентификация плазмид возможна на генетическом и молекулярном уровнях. 1. Генетический уровень Этот метод идентификации основан на учете фенотипов исследуемых бактерий (свежих изолятов) по сравнению с фенотипами известных штаммов, а так же на последующем установлении трансферабельности интересующего свойства к другим бактериям. Что бы проверить, детерминируется ли отличающий признак плазмидой, прибегают к конъюгационным скрещиваниям резистентных клеток (доноры) с чувствительными клетками (акцепторы). 2. Молекулярный уровень Идентификация плазмид в этом случае производится путем выделения, очистки и характеристики плазмидной ДНК, количество которой в плазмидосодержащих клетках составляет около 5% тотальной ДНК клетки. Основная сложность в выделении ДНК плазмид – отделение от хромосомной ДНК. Это достигается с помощью ультрацентрифугирования в градиенте плотности хлористого цезия. В исследованиях используют метод электрофореза, с помощью ферментов-рестриктаз получают физические карты плазмид. Молекулярная масса ДНК плазмиды определяется с помощью определения контурной длины молекулы используя электронную микроскопию.
КЛАССИФИКАЦИЯ ПЛАЗМИД Плазмиды в диких бактериальных популяциях распространены очень широко и их можно выявить у бактерий по всей планете. Плазмиды выявляются во многих видах грамотрицательных анаэробов и аэробов, неспоровых анаэробов, кокков, коккобацилл, грамположительных неспоровых палочковых форм и кокков, споровых палочек и кокков, актиномицетов и родственных форм, микоплазм, спирилл, миксококков, фототрофов, цианобактерий, одноклеточных водорослей, дрожжей, трипаносом и т.д. В 50-е гг. плазмиды R стали классифицировать на fi+ и fi- (по способности ингибировать перенос плазмиды F). Далее стали, в зависимости от различия в пилях, выделять F и I-подобные плазмиды. Современные подходы к классификации плазмид основаны на комплексном учете их генетических свойств. Еще в ранних работах по изучению плазмид было замечено, что существуют факторы, препятствующие конъюгационному переносу плазмид от доноров к реципиентам, содержащим одинаковые и сходные плазмиды. Один из таких факторов – поверхностное исключение: в скрещиваниях плазмида не переходит из клеток доноров в клетки реципиенты, содержащие сходную плазмиду. В результате поверхностного исключения перенос снижается в 10-400 раз по сравнению с нормой. Следующий фактор был открыт в 60-е гг. – летальный зигозиз. Смешивание клеток доноров с клетками реципиентами, добавленными в смесь в значительно меньшем количестве, чем клетки доноры, сопровождается снижением числа жизнеспособных зигот, наследующих донорский генетический материал. Наконец результативность переноса зависит от несовместимости плазмид. В наиболее простом виде несовместимость заключается в том, что при переносе одна из плазмид элиминируется. Если в клетке обе плазмиды сохраняются, то это указывает на их совместимость. Обычно несовместимы те плазмиды, контроль репликации которых одинаков. Рассмотрим поверхностное исключение и летальный зигозиз. Поверхностное исключение (sfx) лучше всего исследовано в случае F-плазмид. Экспериментальные данные свидетельствуют, что это свойство плазмид F детерминируется генами tra S и tra T. Мутации этих генов снижают sfx в 15-20 раз. Изучение роли белка tra T плазмид F показало, что этот белок либо снижает частоту формирования стойких клеточных агрегатов в процессе скрещивания клеток, либо связан с концом f-пили, предупреждая взаимодействие последней с поверхностью клетки реципиента. Белок tra S подавляет запуск конъюгационного метаболизма ДНК. Клетки доноры F+ становятся фенокопиями F- в поздней стационарной фазе развития при культивировании и имеют реципиентную способность. В фенокопиях продукт гена tra T также синтезируется, однако функционально не активен. Поверхностное исключение обнаружено также в случае I-подобных плазмид. Летальный зигозиз может проявляться если в скрещиваниях используются клетки реципиенты F-. Однако, если клетки, используемые в качестве реципиентов, содержат плазмиду F - они имунны к летальному зигозису. МОЛЕКУЛЯРНАЯ И ГЕНЕТИЧЕСКАЯ ОРГАНИЗАЦИЯ ПЛАЗМИД Генетическая организация разных плазмид отличается большим разнообразием, так как среди плазмид, на основе их функциональной специфичности, различают факторы генного переноса, представляющие собой структуры, содержащие лишь гены репликации и переноса, благодаря которым обеспечивается непрерывность поддержания плазмид этого типа и распределение их между дочерними клетками, а так же их трансмиссивность и генетические детерминанты различных свойств. Конъюгационные коинтегративные плазмиды – коинтеграты, состоящие из фактора генного переноса(RTF) и генов, детерминирующих фенотипические свойства бактерий. Каждый из составных компонентов такой плазмиды содержит в своем геноме гены репликации. Неконъюгационные плазмиды - генные детерминанты различных свойств. В бактериальных клетках они лишены способности придавать клеткам свойства генных доноров (они не способны к самостоятельной передаче, но, благодаря наличию генов репликации, стабильно поддерживаются в клетке и передаются дочерним клеткам).
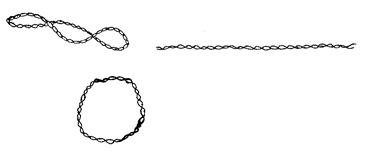
Молекулярная организация Плазмиды – молекулы ДНК, с размерами от 1 Молекуле плазмидной ДНК присущи различные конформации: может быть 2-хцепочечная кольцевая форма (в результате смыкания одной из цепей ДНК – «релаксированная» форма), в результате смыкания обеих цепей образуется ковалентно закрытая сверхспиральная кольцевая форма.
Рисунок 4.1 - Конформации плазмидной ДНК (суперспирализованная, линейная и кольцевая релаксированная)
Для большинства бактерий и плазмид обычна суперспирализированная форма. У микроорганизмов ряда видов встречаются плазмиды в линейной форме, например, у стрептомицетов – плазмида SCP1. Значительная часть сверхспиральной ДНК отдельных плазмид находится в «релаксационном» комплексе с белком. Кольцевая форма молекулы ДНК плазмиды характерна лишь для бактерий, но не для грибов и растений, где она существует в линейной форме.
КЛЕТКАХ Выдающееся свойство плазмид – поддержание в бактериальных клетках в определенном числе копий. Здесь важны процесс репликации и точного распределения между дочерними клетками. У F-плазмиды имеется процесс, контролируемы плазмидой в виде пары генов ccd, определяющий отсеивание клеток, утерявших плазмиду. Заключается в деструкции клеток, в которых произошла ее элиминация. Когда клетка теряет плазмиду, происходит активация продуктов генов ccd, что влечет к запуску механизма самоубийства в клетке и далее в ее потомках.
Генетическая регуляция
Внешнее проявление поддержания плазмид в клетках заключается в том, что большинство или все клетки плазмидосодержащей популяции содержат плазмиды. В случае F, RI и RK2 установлено, что они кодируют летальность клеток, потерявших плазмиду, т.к. продукты соответствующих плазмидных убивают клетки, потерявшие их. Некоторые плазмиды поддерживаются в клетках в количестве 1 – 3 копий на клетку. Они нуждаются для репликации в ДНК-полимеразеIII и их репликация проходит под строгим контролем клетки. Другие – в количестве 40–50 копий и используют ДНК-полимеразу I. Их репликация проходит под релаксированным контролем. При делении клетки, дочерние получают не менее 1 копии плазмиды. Так как считалось, что они реплицируются на определенной стадии репликации ДНК бактерии, то, вероятно, существуют контрольные механизмы. Было предположено, что плазмиды сами регулируют свою репликацию и распределение, причем система репликации реализуется путем осуществления двух функций, одна и которых определяет среднее количество копий на клетку, а другая отзывается на изменение количества и восстанавливает его до нормы. КОНЪЮГАЦИОННЫЙ ПЕРЕНОС Процесс конъюгационного переноса контролируется опероном tra. В общем виде он представляет из себя: 1) образование конъюгационных пар. 2) установление специфичных клеточных контактов. Перенос начинается с oriT. Перенесенная ДНК может стойко поддерживаться в трансконъюгантах в автономном состоянии либо включаться в хромосомный репликон хозяина посредством рекомбинации. Долгое время считалось, что конъюгация – исключительно свойство Enterobacteriaceae, однако в последние годы показана ее возможность и у других микроорганизмов. У Enterobacteriaceae и Pseudomonadaceae конъюгационный перенос обеспечивается тем, что плазмиды контролируют синтез секс-пилей для контактов. У грамположительных бактерий не обнаружены, у некоторых из них контакты между клетками обеспечиваются транспозонами (Streptococcus), фагами (staphylococcus) или экстрахромосомными ферромонами (Enterococcus). При формировании клеточных контактов наряду с парами образуются агрегаты, в которых насчитывают до 13 скрещивающихся клеток. Эффективность спаривания не зависит от размеров агрегатов. Большинство скрещиваемых клеток способно формировать агрегаты в течение 30 минут, что, как предполагают, - результат роста и деления клеток, установивших контакты. Способность спариваться клеток-доноров с клетками-реципиентами определяется наличием F-пилей у доноров. [8, c. 167] Что бы быть компетентной в конъюгации, клетки-реципиенты должны обладать рядом свойств: · Клеточная стенка должна иметь специфические рецепторы. · Клеточная стенка должна пропускать одноцепочечную плазмидную ДНК. Внешние факторы оказывают большое влияние на конъюгацию бактериальных клеток (состав питательной среды, температура, время культивирования, изменение pH среды: снижение pH от 7.2 до 6.2 ведет к удвоению частоты спариваний). Плазмидный перенос состоит из ряда стадий: · Инициация ДНК плазмиды. · Разделение цепей переносимой ДНК в клетке доноре. Для этого необходима насечка «надсечка» и раскручивание макромолекулы, что обеспечивается эндонуклеазами клетки – никазами. · Перенос цепи. Он осуществляется в направлении от 5’ к 3’-концу, т.е. 3’-конец – ведущий. Перенос длится 15 – 20 минут. · Конъюгативный синтез в клетке доноре · Репликонация ДНК плазмиды в клетке-реципиете. Репликонация – конвертирование одноцепочечной ДНК плазмиды в двухцепочечную. Репликонация осуществляется в виде 2-х процессов: синтеза комплиментарной цепи и кольцевания плазмиды. Изучение плазмид F и сol показало необходимость для конъюгационного синтеза в клетках реципиентах ДНК-полимеразы III (для синтеза цепи). ДНК-полимеразе для работы необходима затравка, которую обеспечивает либо РНК-полимераза, либо РНК-примаза. ДНК плазмид прикрепляется ко внутренней мембране в реципиентных клетках, где она приобретает кольцевую форму после синтеза второй цепи. Мобилизация – процесс, основу которого составляют метаболические реакции, обеспечивающие подготовку плазмиды к переносу. Мобилизация присуща как конъюгативным, так и неконъюгативным плазмидам. Конъюгативные плазмиды могут мобилизовать на перенос неконъюгативные плазмиды. Это применяется при скрещивание даже очень отдаленным видам. Механизмами, ограничивающими перенос, являются поверхностное исключение, которое снижает вероятность переноса 100-300 раз и детерминируется плазмидными генами traS traT и неэффективность пилей. Кроме того, существуют ограничения, действующие после проникновения плазмид в клетки, так в результате изменения экспрессии плазмидных генов, клетки иногда не получают донорских свойств, плазмиды могут разрушаться рестриктазами, кодируемыми внутриклеточ-ным генетическим материалом (предполагают, что конъюгативные хромосомы с широким диапазоном переноса обладают генами с антирестрикционными функциями. Важными механизмом является летальный зигозис.
ПЛАЗМИДАМИ a) Плазмиды лекарственной устойчивости. § Общая характеристика и механизмы действия. § Мутации внехромосомных детерминантов резистентности. § Элиминация R-плазмид. § Лекарственная конверсия. § Продление чувствительности к лекарствам. b) Плазмиды бактериоциногении. c) Плазмиды и патогенность бактерий. § Атрибуты патогенности. § Плазмиды и патогенность E. coli. По данным японских исследователей, первый случай выделения бактерий, устойчивых к нескольким лекарственным веществам с различными механизмами действия, был отмечен в 1955 г. в клинике при обследовании женщины, больной дизентерией. От нее была выделена шигелла, устойчивая к действию сульфамидов, стрептомицина, хлорамфеникола и тетрациклина. Вскоре после этого в 1956 г. было выделено большое количество дизентерийных штаммов, обладавших резистентностью к тем же четырем лекарственным веществам, причем во многих случаях лечение проводилось лишь одним из них. Вслед за японскими работами появились сообщения о множественной резистентности, выявленной у других представителей семейства энтеробактерий и стафилококков. Японские ученые доказали возможность передачи всего комплекса устойчивости от его носителей к чувствительным бактериям. Для обозначения комплекса генетических детерминантов множественной лекарственной устойчивости японские исследователи предложили символ R. R-плазмиды детерминируют резистентность бактерий к лекарственным веществам – главным образом антибио-тикам и сульфаниламидам. В широком плане к плазмидам резистентности относят плазмиды, кодирующие резистентность бактерий к бактериофагам, бактерио-цинам, сыворотке, ионизирующему излучению, тяжелым металлам и др. Наиболее полно изучены R-плазмиды грамотрицательных бактерий. Большинство из них конъюгативные, а R-плазмиды грамположительных – неконъюгативные. Большинство генов устойчивости транспозонного происхождения. Их отличительная особенность в том, что они включаются в определенных местах. Такие последовательности именуются интегронами, а их носителями являются обычно транспозоны. ДНК R-плазмид представлена в виде ковалентно закрытых кольцевых молекул, предположительно в суперспирализированной форме.
Таблица 7.1 – Молекулярная масса, копийность и другие свойства некоторых R-плазмид.
Резистентность бактерий, содержащих R-плазмиды, обычно зависит, от вида бактерий и типа антибиотиков. Для плазмидных детерминантов резистентности часто характерно повышенная экспрессивность. Повышение копийности плазмид в результате мутаций генов репликации сопровождается значительным повышением уровня резистентности и подчиняется закономерности «доза-эффект». Повышении экспрессии плазмидных детерминантов лекарственной устойчивости может происходить также и посредством других механизмов: амплификация, приобретение множественных копий транспозонов, повышение интенсивности транскрипции генов. При смешанном культивировании бактерий различной видовой принадлежности возможен межродовой перенос некоторых плазмид.
Таблица 7.2 - Лекарственная резистентность энтеропатогенных штаммов E. coli, выделенных в Англии в 1980-81 гг.
Всего 232 штамма, из них 134 – с резистентностью, а 65 – с трансферрабельной резистентностью. Возрастание частоты резистентности и встречаемости в резистентных штаммах R-плазмид происходит по мере использования антибиотиков и др. Значительный рост резистентности, обусловленный плазмидами, отмечается у стрептококков, причем частота зависит от антибиотика. Возрастание частоты резистентности четко видно на стафилококках (S. aureus) – частых возбудителей внутригоспитальных инфекций. Плазмиды могут участвовать в формировании хромосомной резистентности в качестве векторов транспозируемых генетических элементов. Частота, с которой появляются резистентные бактерии в среде очень высока. Поскольку лекарственные вещества и кормовые антибиотики используются в ветеринарии и растениеводстве, то распространение селекционированных резистентных организмов от одного к другому хозяину, а так же от животных к человеку имеет важное эпизоотическое и эпидемиологическое значение. Ферментативная инактивация антибиотика происходит в среде за пределами клетки. Одиночные клетки поэтому беззащитны. Ферменты b-лактамазы инактивируют b-лактамные антибиотики путем гидролиза еще до того, как они успеют проникнуть через клеточную мембрану и достичь пенициллинсвязывающих белков в ЦПМ. Изменение сайтов-мишеней в качестве механизма резистентности показаны с случае многих антибиотиков. Однако прямое отношение доказано лишь в случае эритромицина и линкомицина. Механизмы резистентности к этим антибиотикам заключаются в специфическом N6-диметилировании 2-х адениновых остатков в рРНК, в результате чего, такие рибосомы в значительно меньшей степени связываются с антибиотиком (такое связывание привело бы к ингибированию белкового синтеза). Efflux-механизм характерен для резистентности к тетрациклинам. Он связан с удалением из клетки этого антибиотика. Не исключено, что пониженное действие тетрациклина связано как с пониженным его восприятием клеткой, так и с efflux-механизмом. Обходные механизмы лежат в основе плазмидной резистентности к сульфонамидам и триметоприму. Плазмиды обеспечивают клетки новым ферментом, заменяющий подавленный и нечувствительный к ингибирующему действию антибиотика. Кроме всего перечисленного плазмиды определяют также и резистентность к тяжелым металлам и др. Лекарственная конверсия. Детерминанты устойчивости вызывают в клетках бактерий изменения, затрагивающие самые разнообразные признаки и известные в литературе как проявление «лекарственной конверсии». Приводятся сведения о более высокой выживаемости бактериальных клеток, инфицированных некоторыми R-факторами, поле УФ-облучения. Лекарственная конверсия может затрагивать также фаготип сальмонелл Плазмиды бактериоциногении. Бактериоцины – вещества, летальные для клеток бактерий. Их названия определяются названиями микроорганизмов-продуцентов. Это термостабильные белки, массой от 10000 до 90000 дальтон. Плазмиды колициногении (col) наиболее изучены. Они содержатся в 20% штаммов E. coli. Колицины подразделяют на группы: A, B, C, D, E, F, G, H, I, J, K, S1, S2, S3, S4, S5, V. Группа E делится на E1, E2, E3 и тд. E1 вносит непоправимые изменения в цитоплазматическую мембрану. E3 разрушительно воздействует на рРНК, E2 вызывает деградацию ДНК. Плазмиды тоже делятся на 2 группы: I (col E1, col E2, col E3, …E4, …E5, …E6, …E7, …E8, ..E9, col N, col K, col A) и II (col Ib, col B, col V) Среди плазмид колициногении встречаются как конъюгативные, так и неконъюгативные. Некоторые способны мобилизовать хромосомные гены. Все плазмиды колициногении постоянно находятся в автономном состоянии. Колицины действуют только на близкие по виду клетки, адсорбируясь на специальных рецепторах на их поверхности. Для того, чтобы убить чувствительную клетку достаточно нескольких молекул колицинов. Бактериоцины продуцируются и другими клетками. Так Bacillus megaterium продуцируют три вида мегатериоцинов: A, B, C. Атрибуты патогенности. § Способность к адгезии (позволяет конкурировать с бактериями-комменсалами за колонизацию эпителиальных поверхностей). Обеспечивается адгезинами и клеточными рецепторами. Бактериальный адгезин – белковая структура, на поверхности клетки бактерии. Он взаимодействует с рецептором соматической клетки. В качестве адгезинов часто выступают обычные пили или фимбрии. § Факторы инвазивности - гемолизины, гиалуро-нидаза, протеаза, ДНК-аза, лецитиназа. § Антифагоцитарные свойства – капсула и др. § Токсины. ЗАКЛЮЧЕНИЕ
Из рассмотренного видно, какую большую роль играют плазмиды в существовании бактерий и, следовательно, человека. Актуальные задачи генетики микроорганизмов в настоящее время невозможно представить без систематического изучения роли и механизмов действия внехромосомных факторов наследственности. Исследование генетической природы, молекулярной структуры и регуляции функций плазмид представляет собой одну из фундаментальных проблем современной биологии. Популярность внехромосомных факторов наследственности микроорганизмов в качестве экспериментальной модели для исследований в разных аспектах объясняется широтой распространения явлений, контролируемых ими и не укладывающихся в рамки хромосомных мутаций, относительной простотой методических приемов идентификации внехромосомных элементов, неограниченной возможностью манипуляций с целью генетического моделирования и четкостью результатов физико-химического анализа их структуры и физиологических функций. Перспектива использования плазмид и эписом в связи с их исключительной биологической пластичностью оценивается молекулярными биологами трезво и объективно. Интенсивно ведется изучение в области генной инженерии. Объективное представление об истинном значении в Представляется оправданным более широкий фронт исследований, направленных на изучение взаимодействия плазмид с целью создания стабильных генетических систем, обеспечивающих более высокую конкурентоспособность (приживляемость в условиях живого организма) обладающих ими бактерий хозяев. Вполне реально конструирование штаммов-продуцентов, несущих целенаправленные сочетания дерепрессированных плазмид со свойствами специфического ингибирования репликации и передачи факторов патогенности. По-видимому, в недалеком будущем на повестку дня станет создание комплексных бакпрепаратов, соответствующих задачам коррекции определенного типа микробиоценоза и дисбактериозов, сопряженных с периодическим доминированием условно патогенных и патогенных микроорганизмов. Такие задачи естественно возникают при анализе замкнутых экосистем, а также при разработке теоретических проблем эволюции микроорганизмов. Таким образом, в данной работе: 1. была изучена история исследования плазмид; 2. были рассмотрены особенности идентификации и классификации плазмид; 3. была рассмотрена генетическая и молекулярная организацию плазмид; 4. были изучены особенности поддержания плазмид в бактериальных клетках; 5. была выявлена сущность конъюгационного переноса; 6. Были рассмотрены свойства бактерий, контролируемые бактериями.
СПИСОК ИСПОЛЬЗОВАННОЙ ЛИТЕРАТУРЫ
1. А.А.Воробьев, А.С.Быков. Микробиология.- М., 1995. – 275 с. 2. Борисов Л.Б. Медицинская микробиология, вирусология, иммунология. - М., 2004. – 313 с. 3. В.Д.Тимаков, В.С. Левашев, Л.Б.Борисов. Микробиология, 1983. – 199 с. 4. Гусев М. В., Минеева Л. А. Микробиология. М.: Медицина, 2003. – 256 с. 5. Канапина А. Ш. «Изучение природы плазмид B. subtilis» Москва, РАН Институт Общей Генетики имени Н. И. Вавилова, 1995 г. – 231 с. 6. Кудлай Д. Г. «Внехромосомные факторы наследственности и их значение в инфекционной патологии» Москва, «Медицина», 1997 г. – 323 с. 7. Пехов А. П. «Основы плазмидологии» Москва, издательство Российского Университета Дружбы Народов, 1996 г. – 311 с. 8. Шабарова З. А., Богданов А. А., Золотухин А. С. «Химические основы генетической инженерии» Москва, издательство МГУ, 1994 г. – 354 с.
ВВЕДЕНИЕ
Плазмиды – внехромосомные генетические элементы, способные к автономному поддержанию в цитоплазме бактерий или существованию в интегрированном в хромосому состоянии, откуда они могут свободно выходить в цитоплазму (иногда с фрагментами хромосомы). Некоторые хромосомы могут распространяться в бактериальной популяции между ее членами. Плазмиды определяют ряд важных свойств бактерий: · Являются факторами фертильности – определяют донорский фенотип клетки. · Контролируют резистентность к антибиотикам, сульфаниламидам, катионам тяжелых металлов, бактериоцинам, бактериофагам, к сыворотке крови. · Чувствительность к бактериоцинам · Синтез тиамина, пролина, внеклеточной ДНКазы и др. · Синтез антибиотиков и бактериоцинов. · Метаболизм углеводов, углеводсодержащих соединений, галогеновых соединений, белков. · Фиксацию азота. · Продукцию токсинов, гемолизина, антигенов колонизации, капсулы. В последнее время природа факторов внехромосомной наследственности микроорганизмов приобрела особый интерес в связи с появлением данных о возможности использования плазмид в качестве векторов эукариотных генов. Такая возможность открывает неограниченные перспективы для генетического моделирования не только при решении проблем молекулярной биологии, но и в практическом аспекте, в частности в медицинской микробиологии и иммунологии (создание новых бактерийных профилактических и лечебных препаратов) и микробиологической промышленности. [3, c. 98] Большой опыт экспериментального мутагенеза на модели бактерий и вирусов способствовал раскрытию генетических и молекулярных механизмов регуляции функций внехромосомных элементов. Их способность включаться в хромосому и формировать комплексы «замещенных» плазмид широко используется в экспериментальной биологии и генетике. Замещенные плазмиды несут фрагменты хромосомы бактерии-хозяина и в автономном состоянии функционируют под контролем регуляторных механизмов бактериальной клетки. Расширение методических и технических возможностей экспериментальных исследований в области молекулярной биологии позволяет целенаправленно использовать генетические модели в решении важных практических задач. Определились реальные пути более гибкого вмешательства в процессы физиологически нормального генетического обмена у бактерий, осуществляемого с участием внехромосомных элементов, способствующих конъюгации, формированию рекомбинантов, передаче генетического материала путем трансдукции умеренными фагами, мобилизации нетрансмиссивных элементов плазмидами, имеющими в своей структуре «гены трансмиссивности», и сочетания с этими генами фрагментов хромосомы с последующим переносом вновь формирующихся структур и их ассоциаций в клетки реципиентов. Актуальное значение приобретает исследование механизмов взаимодействия внехромосомных элементов с хромосомой и между собой в естественных или сконструированных искусственно полиплазмидных системах. Подчинение этих систем общим регуляторным механизмам на уровне клетки и популяции микроорганизмов выдвигает новые проблемы: изучение специфических особенностей полиплазмидных популяций при наличии дополнительных генетических факторов, не обязательных для воспроизведения жизнеспособного потомства, и возможностей практического использования искусственно обогащенного генома популяций бактерий. В последнее десятилетие интенсивно накапливаются данные о генетической природе и биологических особенностях плазмид, с которыми непосредственно связана патологическая активность бактерий. Это – элементы Hly, Ent, Vir, сведения о которых в мало обобщены. Практическое значение в инфекционной патологии приобретают «вторичные» процессы при ожоговых заболеваниях и постхирургических осложнениях, возникающих в связи с неограниченно возрастающей множественностнной лекарственной устойчивостью возбудителей этих процессов, контролируемой трансмиссивными и нетрансмиссивными факторами инфекционной резистентности. Менее полно изучены, но не менее важны плазмиды, контролирующие патогенные свойства стафилококков, стрептококков, псевдомонад. [7, c. 163] Популярное:
|
Последнее изменение этой страницы: 2016-07-13; Просмотров: 891; Нарушение авторского права страницы
 350 МД и более (1000
350 МД и более (1000