
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Адаптация организма к условиям внешней среды и её механизмы. Биологические ритмы организма.
Адаптация - это приспособление строения, функций органов и организма в целом, а также популяции живых существ к изменениям окружающей среды. Различают генотипическую и фенотипическую адаптацию. В основе первой лежат механизмы мутаций, изменчивости, естественного отбора. Они явились причиной формирования современных видов животных и растений. Фенотипическая адаптация - это процесс, протекающий в течение индивидуальной жизни. В результате него организм приобретает устойчивость к какому-либо фактору внешней среды. Это позволяет ему существовать в условиях значительно отличающихся от нормальных. В физиологии и медицине это также процесс сохранения нормального функционального состояния гомеостатических систем, которые обеспечивают развитие, сохранение нормальной работоспособности и жизнедеятельности человека в экстремальных условиях. Выделяют также сложные и перекрестные адаптации. Сложные адаптации возникают в естественных условиях, например к условиям определенных климатических зона, когда организм человека подвергается влиянию комплекса патогенных факторов (на Севере низкая температура, пониженное атмосферное давление, изменение длительности светового дня и т.д.). Перекрестные или кросс-адаптации - это адаптации, при которых развитие устойчивости к одному фактору, повышает резистентность к сопутствующему. Существует два типа адаптивных приспособительных реакций. Первый тип называют пассивным. Эти реакции проявляются на клеточно-тканевом уровне и заключается в формировании определенной степени устойчивости или толернтности к изменениям интенсивности действия какого-либо патогенного фактора внешней среды, например пониженного атмосферного давления. Это позволяет сохранять нормальную физиологическую активность организма при умеренных колебаниях интенсивности данного фактора. Второй тип приспособления - активный. Этот тип заключается в активации специфических адаптивных механизмов. В последнем случае адаптация идет по резистивному типу. Т.е. за счет активного сопротивления воздействию. Если интенсивность воздействия фактора на организм отклоняется от оптимальной величины в ту или иную сторону, но параметры гомеостаза при этом остаются достаточно стабильными, то такие зоны колебаний называется зонами нормы. Имеется две подобных зоны. Одна из них расположена в области недостатка интенсивности фактора, другая в области избытка. Любое смещение интенсивности фактора за пределы зон нормы вызывает перегрузку адаптивных механизмов и нарушению гомеостаза. Поэтому за пределами зон нормы выделяют зоны пессимума В процессе адаптации выделяют два этапа: срочный и долговременный. Первый, начальный, обеспечивает несовершенную адаптацию. Он начинается с момента действия раздражителя и осуществляется на основе имеющихся функциональных механизмов (например, усиление теплопродукции при охлаждении). Долговременный этап адаптации развивается постепенно, в результате длительного или многократного воздействия фактора внешней среды. В его основе лежит многократная активизация механизмов срочной адаптации и постепенное накопление структурных перестроек. Примером долговременной адаптации является изменения механизмов теплообразования и теплоотдачи в холодных климатических условиях. Базисом фенотипической является комплекс последовательных морфофизиологических перестроек, направленных на сохранение постоянства внутренней среды. Основным звеном в механизмах адаптации являются связи физиологических функций с генетическим аппаратом клеток. Под действием экстремального фактора среды происходит увеличение нагрузки на функциональную систему. Это ведет к усилению синтеза нуклеиновых кислот и белков в клетках органов, входящих в систему. В результате в них формируется структурный след адаптации. Активизируются аппараты этих клеток, выполняющие базисные функции: энергетический обмен, трансмембранный транспорт, сигнализацию. Именно этот структурный след является основой долговременной фенотипической адаптации. Однако адаптационные механизмы позволяют компенсировать изменения фактора среды лишь в определенных пределах и определенное время. В результате воздействия на организм факторов, превышающих возможности адаптационных механизмов, развивается дизадаптация. Она приводит к дисфункции систем организма. Следовательно, происходит переход адаптационной реакции в патологическую - болезнь. Примером болезней дизадаптации являются сердечно-сосудистые заболевания у не коренных жителей Севера.
Виды сокращения скелетных мышц. Одиночные мышечные сокращения и его фазы. Суммация мышечных сокращений. Зубчатый и гладкий тетанус. Оптимум и пессимум раздражения (Н.В.Введеннский) При нанесении на двигательный нерв или мышцу одиночного порогового или сверхпорогового раздражения, возникает одиночное сокращение. При его графической регистрации, на полученной кривой можно выделить три последовательных периода: 1. Латентный период. Это время от момента нанесения раздражения до начала сокращения. Его длительность около 1-2 мс. Во время латентного периода генерируется и распространяется ПД, происходит высвобождение кальция из СР, взаимодействие актина с миозином и т.д. 2. Период укорочения. В зависимости от типа мышцы (быстрая или медленная) его продолжительность от 10 до 100 мсек., 3. Период расслабления. Его длительность несколько больше, чем укорочения. В режиме одиночного сокращения мышца способна работать длительное время без утомления, но его сила незначительна. Поэтому в организме такие сокращения встречаются редко, например так могут сокращаться быстрые глазодвигательные мышцы, мышцы сгибателей пальцев. Чаще одиночные сокращения суммируются. Суммация – это сложение двух последовательных сокращений при нанесении на нее двух пороговых или сверхпороговых раздражений, интервал между которыми меньше длительности одиночного сокращения, но больше продолжительности рефракторного периода. Различают 2 вида суммации: полную и неполную суммацию. Неполная суммация возникает в том случае, если повторное раздражение наносится на мышцу, когда она уже начала расслабляться. Полная возникает тогда, когда повторное раздражение действует на мышцу до начала периода расслабления, т.е. в конце периода укорочения. Амплитуда сокращения при полной суммации выше, чем неполной. Если интервал между двумя раздражениями еще больше уменьшить, например, нанести второе в середине периода укорочения, то суммации не будет, потому что мышца находится в состоянии рефрактерности. Тетанус – это длительное сокращение мышцы, возникающее в результате суммации нескольких одиночных сокращений, развивающихся при нанесении на нее ряда последовательных раздражений. Различают 2 формы тетануса: зубчатый и гладкий. Зубчатый тетанус наблюдается в том случае, если каждое последующее раздражение действует на мышцу, когда она уже начала расслабляться. Т.е. наблюдается неполная суммация. Гладкий тетанус возникает тогда, когда каждое последующее раздражение наносится в конце периода укорочения. Т.е. имеет место полная суммация отдельных сокращений. Амплитуда гладкого тетануса больше, чем зубчатого. В норме мышцы человека сокращаются в режиме гладкого тетануса. Зубчатый возникает при патологии, например, тремор рук при алкогольной интоксикации и болезни Паркинсона. Механизм возникновения потенциала действия. Движение ионов натрия и калия при возбуждении. Опыты, доказывающие значение ионов натрия в происхождении потенциала действия. Понятие об электромеханическом сопровождении. Дальнейшие исследования Ходжкина и Хаксли показали, что при возбуждении аксона кальмара возникает быстрое колебание мембранного потенциала, которое на экране осциллографа имело форму пика (spike). Они назвали это колебание потенциалом действия (ПД). Так как электрический ток для возбудимых мембран является адекватным раздражителем, ПД можно вызвать, поместив на наружную поверхность мембраны отрицательный электрод – катод, а внутреннюю положительный - анод. Это приведет к снижению величины заряда мембраны – ее деполяризации. При действии слабого допорогового тока происходит пассивная деполяризация, т.е. возникает катэлектротон. Если силу тока увеличить до определенного предела, то в конце периода его воздействия на плато катэлектротона появится небольшой самопроизвольный подъём – местный или локальный ответ. Он является следствием открывания небольшой части натриевых каналов, находящихся под катодом. При токе пороговой силы МП снижается до критического уровня деполяризации (КУД), при котором начинается генерация потенциала действия. Он находится для нейронов примерно на уровне – 50 мВ.
1. Локальный ответ (местная деполяризация), предшествующий развитию ПД. 2. Фаза деполяризации. Во время этой фазы МП быстро уменьшается и достигает нулевого уровня. Уровень деполяризации растет выше нуля. Поэтому мембрана приобретает противоположный заряд – внутри она становится положительной, а снаружи отрицательной. Явление смены заряда мембраны называется реверсией мембранного потенциала. Продолжительность этой фазы у нервных и мышечных клеток 1-2 мс. 3. Фаза реполяризации. Она начинается при достижении определенного уровня МП (примерно +20 мВ). Мембранный потенциал начинает быстро возвращаться к потенциалу покоя. Длительность фазы 3-5 мс. 4. Фаза следовой деполяризации или следового отрицательного потенциала. Период, когда возвращение МП к потенциалу покоя временно задерживается. Он длится 15-30 мс. 5. Фаза следовой гиперполяризации или следового положительного потенциала. В эту фазу МП на некоторое время становится выше исходного уровня ПП. Ее длительность 250-300 мс. Амплитуда ПД скелетных мышц в среднем 120-130 мВ, нейронов 80-90 мВ, гладкомышечных клеток 40-50 мВ. При возбуждении нейронов ПД возникает в начальном сегменте аксона – аксонном холмике. Возникновение ПД обусловлено изменением ионной проницаемости мембраны при возбуждении. В период локального ответа открываются медленные натриевые каналы, а быстрые остаются закрытыми, возникает временная самопроизвольная деполяризация. Когда МП достигает критического уровня, закрытые активационные ворота натриевых каналов открываются и ионы натрия лавинообразно устремляются в клетку, вызывая нарастающую деполяризацию. В эту фазу открываются и быстрые, и медленные натриевые каналы. Т.е. натриевая проницаемость мембраны резко возрастает. Причем от чувствительности активационных ворот зависит величина КУД (чем она выше, тем ниже КУД, и наоборот). Когда величина деполяризация приближается к равновесному потенциалу для ионов натрия (+20 мВ), сила концентрационного градиента натрия значительно уменьшается. Одновременно начинается процесс инактивации быстрых натриевых каналов и снижения натриевой проводимости мембраны. Деполяризация прекращается. Резко усиливается выход ионов калия, т.е. калиевый выходящий ток. В некоторых клетках это происходит из-за активации специальных каналов калиевого выходящего тока. Этот ток, направленный из клетки, служит для быстрого смещения МП к уровню потенциала покоя. Т.е. начинается фаза реполяризации. Возрастание МП приводит к закрыванию и активационных ворот натриевых каналов, что еще больше снижает натриевую проницаемость мембраны и ускоряет реполяризацию. Возникновение фазы следовой деполяризации объясняется тем, что небольшая часть медленных натриевых каналов остается открытой. Следовая гиперполяризация связана с повышенной после генерации ПД калиевой проводимостью мембраны и тем, что более активно работает натрий-калиевый насос, выносящий вошедшие в клетку во время ПД ионы натрия. Изменяя проводимость быстрых натриевых и калиевых каналов, можно влиять на генерацию ПД, а, следовательно, на возбуждение клеток. При полной блокаде натриевых каналов, например, ядом рыбы тетродонта – тетродотоксином, клетка становится невозбудимой. Это используется в клинике. Такие местные анестетики, как новокаин, дикаин, лидокаин тормозят переход натриевых каналов нервных волокон в открытое состояние. Поэтому проведение нервных импульсов по чувствительным нервам прекращается, наступает обезболивание (анестезия) органа. При блокаде калиевых каналов затрудняется выход ионов калия из цитоплазмы на наружную поверхность мембраны, т.е. восстановление МП. Поэтому удлиняется фаза реполяризации. Этот эффект блокаторов калиевых каналов также используется в клинической практике. Например, один из них хинидин, удлиняя фазу реполяризации кардиомиоцитов, урежает сердечные сокращения и нормализует сердечный ритм. Также следует отметить, что чем выше скорость распространения ПД по мембране клетки или ткани, тем выше ее проводимость.
Передача сигнала от возбужденной мембраны к миофибриллам называется электромеханическим сопряжением. Когда генерация ПД прекращается и мембранный потенциал возвращается к исходному уровню, начинает работать Са-насос (фермент Са-АТФаза). Ионы кальция вновь закачиваются в цистерны саркоплазматического ретикулума и их концентрация падает ниже 10-8 моль. Молекулы тропонина приобретают исходную форму и тропомиозин вновь начинает блокировать активные центры актина. Головки миозина отсоединяются от них и мышца за счет эластичности приходит в исходное расслабленное состояние. Популярное:
|
Последнее изменение этой страницы: 2016-07-13; Просмотров: 2810; Нарушение авторского права страницы
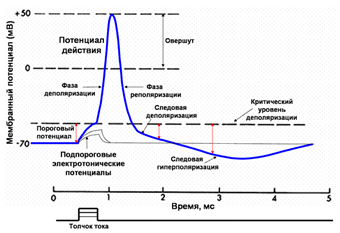 На кривой потенциала действия выделяют следующие фазы:
На кривой потенциала действия выделяют следующие фазы: