|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
КАФЕДРА БИОЛОГИИ С ЭКОЛОГИЕЙСтр 1 из 6Следующая ⇒
ГОСУДАРСТВЕННОЕ ОБРАЗОВАТЕЛЬНОЕ УЧРЕЖДЕНИЕ ВЫСШЕГО ПРОФЕССИОНАЛЬНОГО ОБРАЗОВАНИЯ «СТАВРОПОЛЬСКАЯ ГОСУДАРСТВЕННАЯ МЕДИЦИНСКАЯ АКАДЕМИЯ ФЕДЕРАЛЬНОГО АГЕНСТВА ПО ЗДРАВООХРАНЕНИЮ И СОЦИАЛЬНОМУ РАЗВИТИЮ» КАФЕДРА БИОЛОГИИ С ЭКОЛОГИЕЙ ХОДЖАЯН А.Б., МИХАЙЛЕНКО А.К., МАКАРЕНКО Э.Н. Основы ЦИТОЛОГИИ:
СТРУКТУРНАЯ ОРГАНИЗАЦИЯ КЛЕТКИ
Учебное пособие для студентов первого курса ФВСО
Ставрополь, 2009 УДК: 576.3 (07) ББК 28.05я73 Х 69 «Основы цитологии: структурная организация клетки», Учебное пособие для студентов первого курса факультета высшего сестринского образования (ФВСО). – Ставрополь: Изд - во СтГМА. – 2009. – 50 с. Авторы: Ходжаян Анна Борисовна, доктор медицинских наук, профессор, заведующая кафедрой биологии с экологией; Михайленко Антонина Кузьминична, кандидат биологических наук, старший преподаватель кафедры биологии с экологией; Макаренко Элина Николаевна, кандидат медицинских наук, старший преподаватель кафедры биологии с экологией. Учебное пособие включает необходимые материалы по разделу «Основы цитологии: структурная организация клетки» для самоподготовки студентов ФВСО заочной формы обучения. Данное пособие дополняет основную учебную литературу, содержит данные, не вошедшие в учебники, некоторые новые современные положения, оригинальный иллюстративный материал, контрольные тестовые вопросы и способствует более глубокой проработке изучаемого предмета. Рецензенты: Мещеряков А.Ф., академик РАЕН, академик МАО, доктор биологических наук, профессор. Чижова Л.Н., зав. лабораторией иммуногенетики, биохимии, общей химии ГНУ СНИИЖК, доктор сельскохозяйственных наук, профессор УДК: 576.3 (07) ББК 28.05я73 Х 69 Рекомендовано к изданию цикловой методической комиссией Ставропольской государственной медицинской академии.
Рекомендовано к печати редакционно-издательским советом СтГМА.
© Ставропольская государственная медицинская академия, 2009
Краткая история цитологии Цитология (греч. citos – клетка, logos – наука) – наука о клетке. В настоящее время учение о клетке является во многих отношениях центральным объектом биологических исследований. Предпосылкой для открытия клетки явилось изобретение микроскопа и его использование для исследования биологических объектов. Первый световой микроскоп сконструировали в Голландии в 1590 году два брата, Ганс и Захариус Янссены, шлифовальщики линз. Долгое время микроскоп использовался как забава, игрушка для развлечения знатных особ.
Термин «клетка» утвердился в биологии, несмотря на то, что Роберт Гук наблюдал, в действительности, не клетки, а только целлюлозные оболочки растительных клеток. Кроме того, клетки не являются полостями. В дальнейшем клеточное строение многих частей растений видели и описали М. Мальпиги, Н. Грю, а также А. Левенгук. Важным событием в развитии представлений о клетке была изданная в 1672 году книга Марчелло Мальпиги «Анатомия растений», где приводилось подробное описание микроскопических растительных структур. В своих исследованиях Мальпиги убедился, что растения состоят из клеток, которые он называл «мешочками» и «пузырьками». Среди блестящей плеяды микроскопистов XVII века одно из первых мест занимает А. Левенгук, голландский купец, который завоевал себе славу учёного. Он прославился созданием линз, которые давали увеличение в 100-300 раз. В 1674 году Антонио ван Левенгук открыл с помощью собственноручно изобретенного микроскопа одноклеточных простейших, названных им «микроскопическими животными», бактерии, дрожжи, клетки крови – эритроциты, половые клетки – сперматозоиды, которые Левенгук называл «анималькули». Из животных тканей Левенгук изучал и точно описал строение сердечной мышцы. Он был первым натуралистом, наблюдавшим клетки животногоорганизма. Это пробудило интерес к изучению живого микромира. Как наука цитология возникла лишь в XIX веке. В это время были сделаны важные открытия. В 1830 году чешский исследователь Ян Пуркинье описал вязкое студенистое вещество внутри клетки и назвал его протоплазмой (гр. protos – первый, plasma – образование). В 1831 году шотландский ученый Роберт Броун открыл ядро. В 1836 году Габриелем Валентини в ядре было обнаружено ядрышко.
В 1838 году была опубликована работа Матиаса Шлейдена «Данные о фитогенезисе», где автор, опираясь на уже имевшиеся в ботанике представления о клетке, выдвинул идею об идентичности растительных клеток с точки зрения их развития. Он пришёл к выводу, что закон клеточного строения справедлив для растений. В 1839 году вышла в свет ставшая классической книга Теодора Шванна «Микроскопические исследования о соответствии в структуре и росте животных и растений». В ней автор сделал окончательный вывод о том, что клетка является структурной единицей жизнедеятельности и развития растений и животных. В 1838 – 1839 годах немецкие ученые Матиас Шлейден и ТеодорШванн независимо друг от друга сформулировали клеточную теорию.
КЛЕТОЧНАЯ ТЕОРИЯ: 1) все живые организмы (растения и животные) состоят из клеток; 2) клетки растений и животных сходны по строению, химическому составу и выполняемым функциям.
М. Шлейден и Т. Шванн считали, что клетки в организме возникают путём новообразования из первичного неклеточного вещества. В 1858 году немецкий учёный-анатом Рудольф Вирхов в своей книге «Целлюлярная патология» опроверг это представление и доказал, что новые клетки всегда возникают из предшествующих путем деления – «клетка от клетки, всё живое только из клетки» – (omnis cellula a cellula). Важным обобщением Р.Вирхова явилось утверждение, что наибольшее значение в жизнедеятельности клеток имеют не оболочки, а их содержимое – протоплазма и ядро. Опираясь на клеточную теорию, Р. Вирхов поставил на научную основу учение о болезнях. Опровергнув господствующее в то время представление, согласно которому в основе болезней лежит только изменение состава жидкостей организма (крови, лимфы, желчи), он доказал огромное значение изменений, происходящих в клетках и тканях. Р. Вирхов установил: «Всякое болезненное изменение связано с каким-то патологическим процессом в клетках, составляющих организм». Это утверждение стало основой для появления важнейшего раздела современной медицины – патологической анатомии. Вирхов был одним из основоположников исследования явлений жизнедеятельности на клеточном уровне, что является его бесспорной заслугой. Однако при этом он недооценивал исследования тех же явлений на уровне организма как целостной системы. В представлении Вирхова организм – это государство клеток и все его функции сводятся к сумме свойств отдельных клеток. В преодолении этих односторонних представлений об организме большое значение имели работы И.М.Сеченова, С.П.Боткина и И.П.Павлова. Отечественные ученые доказали, что организм представляет собой по отношению к клеткам высшее единство. Клетки и другие структурные элементы, составляющие тело, не обладают физиологической самостоятельностью. Их формирование и функции координируются и управляются целостным организмом с помощью сложной системы химической и нервной регуляции. Коренное улучшение всей техники микроскопирования позволило исследователям к началу XX столетия обнаружить основные клеточные органоиды, выяснить строение ядра и закономерности клеточного деления, расшифровать механизмы оплодотворения и созревания половых клеток. В 1876 году Эдуард Ван Бенеден установил наличие клеточного центра в делящихся половых клетках. В 1890 году Рихард Альтман описал митохондрии, назвав их биобластами, и выдвинул идею о возможности их самовоспроизведения. В 1898 году Камилло Гольджи открыл органоид, названный в его честь комплексом Гольджи. В 1898 году хромосомы впервые были описаны Карлом Бенда. Крупный вклад в развитие учения о клетке во второй половине XIX – начале XX вв. внесли отечественные цитологи И.Д.Чистяков (описание фаз митотического деления), И.Н.Горожанкин (изучение цитологических основ оплодотворения у растений), С.Г.Навашин, открывший в 1898г. явление двойного оплодотворения у растений. Успехи в изучении клетки привели к тому, что внимание биологов все больше концентрировалось на клетке как основной структурной единице живых организмов. Качественный скачок в цитологии произошел в XX веке. В 1932 году МаксКнолль и Эрнст Руска изобрели электронный микроскоп, дающий увеличение в 106 раз. Были обнаружены и описаны невидимые в световой микроскоп микро- и ультрамикроструктуры клетки. С этого момента клетку начали изучать на молекулярном уровне. Таким образом, достижения цитологии всегда связаны с усовершенствованием техники микроскопирования.
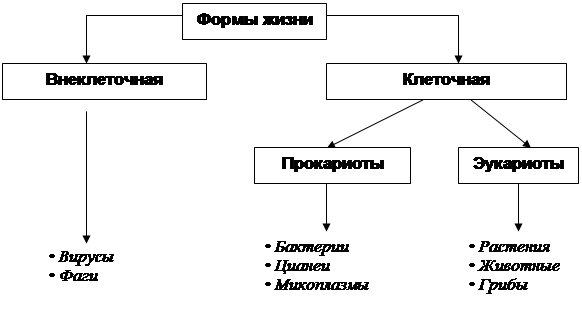
ФОРМЫ ЖИЗНИ Жизнь на планете Земля известна только в двух формах: внеклеточной и клеточной. Внеклеточная форма жизни – это особая форма, представленная вирусами и бактериофагами (фагами), которые занимают промежуточное положение между живой и неживой природой. Клеточная форма жизни (организмы) в зависимости от типа организации клеток подразделяется на прокариоты и эукариоты. Прокариоты – это одноклеточные организмы, не имеющие оформленного ядра. К ним относятся бактерии, цианеи (цианобактерии или сине-зеленые водоросли) и микоплазмы, образующие царство Дробянки. Эукариоты – это одноклеточные и многоклеточные организмы. В их клетках всегда есть четко оформленное ядро.
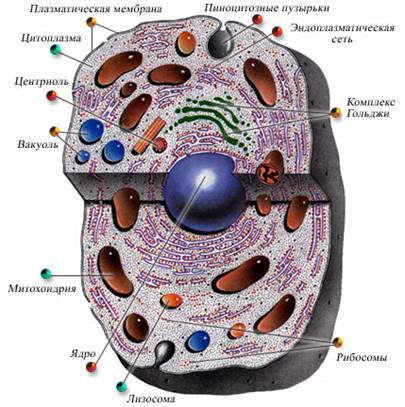
ТИПЫ ОРГАНИЗАЦИИ КЛЕТОК Существуют два типа организации клеток: прокариотические и эукариотические клетки (таблица 1). Принципиальным отличием между ними является наличие оформленного ядра. Прокариотические клетки (доядерные) не содержат оформленного ядра. Основными компонентами прокариотических клеток являются оболочка и цитоплазма. Структурой, отвечающей за передачу наследственной информации, является генофор (нуклеоид), расположенный непосредственно в цитоплазме. По химической природе генофор – это молекула ДНК, не связанная с белками и имеющая форму кольца. На поверхности плазматической мембраны бактерий располагается клеточная стенка, состоящая из муреина (полисахаридных цепей, соединенных друг с другом короткими цепями пептидов). У некоторых клеток клеточная стенка покрыта защитным слизистым слоем или капсулой. Клеточная стенка сохраняет форму клеток, обеспечивает их жесткость и антигенные свойства. Мембрана прокариот имеет сложнодифференцированные впячивания – мезосомы, которые по своим функциям напоминают митохондрии эукариотических клеток. Цитоплазма прокариотических клеток лишена органоидов, за исключением рибосом. В ней находятся включения в виде гранул гликогена, липидов и т.д. Кроме того, в ней присутствуют плазмиды (внекольцевые фрагменты ДНК, определяющие ряд признаков и свойств данной клетки, гены которых контролируют незначительную часть наследственных признаков бактериальной клетки). Они способны к самостоятельной репликации и стабильно наследуются потомством. Широко используются в генной инженерии. У зеленых и пурпурных бактерий (автотрофы) на впячиваниях плазматической мембраны находятся фотосинтезирующие пигменты. Следовательно, клетки сине-зеленых водорослей (цианеи) сходны с бактериальными, но, кроме вышеперечисленных компонентов, они содержат хлорофилл. Эукариотические клетки (ядерные) имеют оформленное ядро, которое включает структуры, ответственные за хранение, воспроизведение и передачу наследственной информации – хромосомы. Они расположены в ядре клетки и отграничены от цитоплазмы ядерной оболочкой. По химической природе хромосомы – это дезоксирибонуклеопротеидные структуры ( ДНП ) – комплексы ДНК и белков. В не делящейся клетке хромосомы деспирализованы и имеют вид нитей хроматина. Во время деления они спирализуются и приобретают палочковидную форму. К надцарству эукариот относятся царства растений, грибов и животных. Для растительных клеток характерно наличие толстой целлюлозной клеточной стенки, различных пластид, крупной центральной вакуоли, смещающей ядро к периферии. В качестве питательного резервного углевода клетки растений запасают крахмал. В клетках грибов клеточная оболочка содержит хитин, в цитоплазме имеется центральная вакуоль, пластиды отсутствуют. Главным резервным полисахаридом является гликоген. Животные клетки имеют цитоплазматическую мембрану, не содержат пластид и центральной вакуоли, для клеточного центра характерны центриоли. Запасным углеводом является гликоген.
Таблица 1 Ядро ЦИТОПЛАЗМАТИЧЕСКАЯ МЕМБРАНА
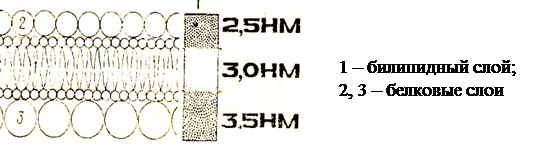
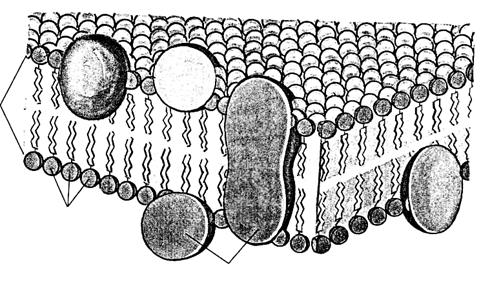
Цитоплазматическая мембрана или плазмалемма (лат. membrana – кожица, плёнка) – тончайшая пленка ( 7– 10нм), отграничивающая внутреннее содержимое клетки от окружающей среды, видна только в электронный микроскоп. По химической организации плазмалемма представляет липопротеидный комплекс – молекулы липидов и белков. Её основу составляет липидный бислой, состоящий из фосфолипидов, кроме этого, в мембранах присутствуют гликолипиды и холестерол. Все они обладают свойством амфипатричности, т.е. у них есть гидрофильные («любящие воду») и гидрофобные («боящиеся воды») концы. Гидрофильные полярные «головки» липидных молекул (фосфатная группа) обращены кнаружи мембраны, а гидрофобные неполярные «хвосты» (остатки жирных кислот) – друг к другу, что создает биполярный липидный слой. Молекулы липидов подвижны и могут перемещаться в своем монослое или редко – из одного монослоя в другой. Монослои липидов обладают ассиметричностью, т. е. отличаются по составу липидов, что придает специфичность мембранам даже в пределах одной клетки. Бислой липидов может находиться в состоянии жидкого или твердого кристалла. Вторым обязательным компонентом плазмалеммы являются белки. Многие мембранные белки способны перемещаться в плоскости мембраны или вращаться вокруг своей оси, но не могут переходить с одной стороны бислоя липидов на другой. Липиды обеспечивают основные структурные особенности мембраны, а белки – её функции. Функции мембранных белков различны: поддержание структуры мембран, получение и преобразование сигналов из окружающей среды, транспорт некоторых веществ, катализ реакций, происходящих на мембранах. Различают несколько моделей строения цитоплазматической мембраны. ①. БУТЕРБРОДНАЯ МОДЕЛЬ (белки – липиды – белки) В 1935г. английские ученые Даниэли и Даусон высказали идею о послойном расположении в мембранемолекул белков (темные слои в электронном микроскопе), которые залегают снаружи, и молекул липидов (светлый слой) – внутри. Длительное время существовало представление о едином трехслойном строении всех биологических мембран. При детальном изучении мембраны с помощью электронного микроскопа оказалось, что светлый слой на самом деле представлен двумя слоями фосфолипидов – это билипидный слой, причем водорастворимые его участки – гидрофильные головки направлены к белковому слою, а нерастворимые (остатки жирных кислот) – гидрофобные хвосты обращены друг к другу.
Однако уже с середины 60-х годов начали накапливаться факты против унитарной «бутербродной» модели. В частности, по одним данным, не все мембраны имели четкую трехслойную структуру при электронно-микроскопическом исследовании; по другим – значительная часть мембранных белков имела глобулярную структуру, а не ламеллярную, как в постулируемой модели. Наконец, среди многочисленных моделей мембран, предложенных в середине 60-х годов, начали выделяться те, в которых доказывалось наличие гидрофобно-гидрофильных взаимодействий не только между липидными молекулами, но и между липидами и белками.
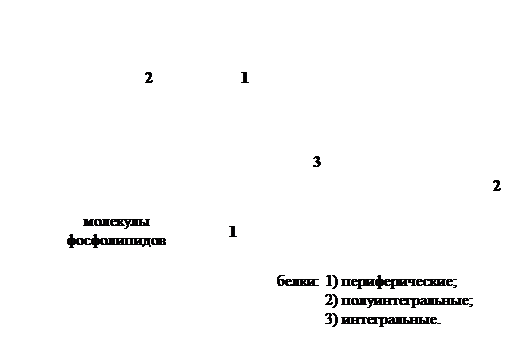
В 1972г. Сингер и Николсон описали модель мембраны, которая получила широкое признание. Согласно этой модели молекулы белков не образуют сплошного слоя, а погружены в биполярный липидный слой на разную глубину в виде мозаики. Глобулы белковых молекул, подобно айсбергам, погружены в «океан»
липидов: одни находятся на поверхности билипидного слоя – периферические белки, другие погружаются в него наполовину – полуинтегральные белки, третьи – интегральные белки – пронизывают его насквозь, формируя гидрофильные поры. Периферические белки, находясь на поверхности билипидного слоя, связаны с головками липидных молекул электростатическими взаимодействиями. Но они никогда не образуют сплошного слоя и, по сути дела, не являются белками собственно мембраны, а, скорее, связывают ее с надмембранной или субмембранной системой поверхностного аппарата клетки. Основную роль в организации собственно мембраны играют интегральные и полуинтегральные (трансмембранные) белки, имеющие глобулярную структуру и связанные с липидной фазой гидрофильно-гидрофобными взаимодействиями. Молекулы белков, как и липиды, обладают амфипатричностью и своими гидрофобными участками взаимодействуют с гидрофобными хвостами билипидного слоя, а гидрофильные участки обращены к водной среде и образуют с водой водородные связи. ③. БЕЛКОВО-КРИСТАЛЛИЧЕСКАЯ МОДЕЛЬ (модель липопротеинового коврика) Мембраны образованы переплетением липидных и белковых молекул, объединяющихся между собой на основе гидрофильно- гидрофобных взаимодействий.
Белковые молекулы, как штифты, пронизывают слой липидов и выполняют в составе мембраны функцию каркаса. После обработки мембраны жирорастворимыми веществами белковый каркас сохраняется, что доказывает взаимосвязь между молекулами белков в мембране. По-видимому, эта модель реализуется лишь в отдельных специальных участках некоторых мембран, где требуется жесткая структура и тесные стабильные взаимоотношения между липидами и белками (например, в области расположения фермента Na-К –АТФ-азы). Самой универсальной моделью, отвечающей термодинамическим принципам (принципам гидрофильно-гидрофобных взаимодействий), морфо-биохимическим и экспериментально-цитологическим данным, является жидкостно-мозаичная модель. Однако все три модели мембран не исключают друг друга и могут встречаться в разных участках одной и той же мембраны в зависимости от функциональных особенностей данного участка. СВОЙСТВА МЕМБРАНЫ 1. Способность к самосборке. После разрушающих воздействий мембрана способна восстановить свою структуру, т.к. молекулы липидов на основе своих физико-химических свойств собираются в биполярный слой, в который затем встраиваются молекулы белков. 2. Текучесть. Мембрана не является жесткой структурой, большая часть входящих в её состав белков и липидов может перемещаться в плоскости мембраны, они постоянно флюктуируют за счет вращательных и колебательных движений. Это определяет большую скорость протекания химических реакций на мембране. 3. Полупроницаемость. Мембраны живых клеток пропускают, помимо воды, лишь определённые молекулы и ионы растворённых веществ. Это обеспечивает поддержание ионного и молекулярного состава клетки. 4. Мембрана не имеет свободных концов. Она всегда замыкается в пузырьки. 5. Асимметричность. Состав наружного и внутреннего слоев как белков, так и липидов различен. 6. Полярность. Внешняя сторона мембраны несёт положительный заряд, а внутренняя – отрицательный.
ФУНКЦИИ МЕМБРАНЫ 1) Барьерная – плазмалемма отграничивает цитоплазму и ядро от внешней среды. Кроме того, мембрана делит внутреннее содержимое клетки на отсеки (компартменты), в которых зачастую протекают противоположные биохимические реакции. 2) Рецепторная (сигнальная) – благодаря важному свойству белковых молекул – денатурации, мембрана способна улавливать различные изменения в окружающей среде. Так, при воздействии на мембрану клетки различных средовых факторов (физических, химических, биологических) белки, входящие в ее состав, меняют свою пространственную конфигурацию, что служит своеобразным сигналом для клетки. Это обеспечивает связь с внешней средой, распознавание клеток и их ориентацию при формировании тканей и т.д. С этой функцией связана деятельность различных регуляторных систем и формирование иммунного ответа. 3) Обменная – в состав мембраны входят не только структурные белки, которые образуют ее, но и ферментативные, являющиеся биологическими катализаторами. Они располагаются на мембране в виде «каталитического конвейера» и определяют интенсивность и направленность реакций метаболизма. 4) Транспортная – молекулы веществ, диаметр которых не превышает 50 нм, могут проникать путем пассивного и активного транспорта через поры в структуре мембраны. Крупные вещества попадают в клетку путем эндоцитоза (транспорт в мембранной упаковке), требующего затраты энергии. Его разновидностями являются фаго- и пиноцитоз. Пассивный транспорт – вид транспорта, в котором перенос веществ осуществляется по градиенту химической или электрохимической концентрации без затраты энергии АТФ. Выделяют два вида пассивного транспорта: простая и облегченная диффузия. Диффузия – это перенос ионов или молекул из зоны более высокой их концентрации в зону более низкой концентрации, т.е. по градиенту. Простая диффузия – ионы солей и вода проникают через трансмембранные белки или жирорастворимые вещества по градиенту концентрации. Облегченная диффузия – специфические белки-переносчики связывают вещество и переносят его через мембрану по принципу «пинг-понга». Таким способом через мембрану проходят сахара и аминокислоты. Скорость такого транспорта значительно выше, чем простой диффузии. Кроме белков- переносчиков, в облегченной диффузии принимают участие некоторые антибиотики – например, грамитидин и ваномицин. Поскольку они обеспечивают транспорт ионов, их называют ионофорами. Активный транспорт – это вид транспорта, при котором расходуется энергия АТФ, он идёт против градиента концентрации. В нем принимают участие ферменты АТФ-азы. В наружной клеточной мембране находятся АТФ-азы, которые осуществляют перенос ионов против градиента концентрации, это явление называется ионным насосом. Примером является натрий-калиевый насос. В норме в клетке больше ионов калия, во внешней среде – ионов натрия. Поэтому по законам простой диффузии калий стремится из клетки, а натрий – в клетку. В противовес этому натрий-калиевый насос накачивает против градиента концентрации в клетку ионы калия, а ионы натрия выносит во внешнюю среду. Это позволяет поддерживать постоянство ионного состава в клетке и её жизнеспособность. В животной клетке одна треть АТФ расходуется на работу натрий-калиевого насоса. Разновидностью активного транспорта является транспорт в мембранной упаковке – эндоцитоз. Крупные молекулы биополимеров не могут проникать через мембрану, они поступают в клетку в мембранной упаковке. Различают фагоцитоз и пиноцитоз. Фагоцитоз – захват клеткой твердых частиц, пиноцитоз – жидких частиц. В этих процессах выделяют стадии: 1) узнавание рецепторами мембраны вещества; 2) впячивание (инвагинация) мембраны с образованием везикулы (пузырька); 3) отрыв пузырька от мембраны, слияние его с первичной лизосомой и восстановление целостности мембраны; 4) выделение непереваренного материала из клетки (экзоцитоз). Эндоцитоз является способом питания для простейших. У млекопитающих и человека имеется ретикуло-гистио-эндотелиальная система клеток, способная к эндоцитозу – это лейкоциты, макрофаги, клетки Купфера в печени.
ОСМОТИЧЕСКИЕ СВОЙСТВА КЛЕТКИ Осмос – односторонний процесс проникновения воды через полупроницаемую мембрану из области с меньшей концентрацией раствора в область с более высокой концентрацией. Осмос обусловливает осмотическое давление. Диализ – односторонняя диффузия растворенных веществ. Раствор, в котором осмотическое давление такое же, как и в клетках, называют изотоническим. При погружении клетки в изотонический раствор её объем не изменяется. Изотонический раствор называют физиологическим – это 0, 9% раствор хлорида натрия, который широко применяется в медицине при сильном обезвоживании и потери плазмы крови. Раствор, осмотическое давление которого выше, чем в клетках, называют гипертоническим. Клетки в гипертоническом растворе теряют воду и сморщиваются. Гипертонические растворы широко применяются в медицине. Марлевая повязка, смоченная в гипертоническом растворе, хорошо впитывает гной. Раствор, где концентрация солей ниже, чем в клетке, называют гипотоническим. При погружении клетки в такой раствор вода устремляется в нее. Клетка набухает, ее тургор увеличивается, и она может разрушиться. Гемолиз – разрушение клеток крови в гипотоническом растворе. Осмотическое давление в организме человека в целом регулируется системой органов выделения.
Типы ЭПС
КОМПЛЕКС (АППАРАТ) ГОЛЬДЖИ Органоид общего значения, имеющий одномембранный принцип строения. В 1898 году итальянский ученый Камилло Гольджи выявил и описал в нервных клетках образования, которые он назвал «внутренним сетчатым аппаратом». Структурной и функциональной единицей комплекса Гольджи является диктиосома. В растительной клетке количество диктиосом больше, чем в животной. На одну диктиосому приходятся4–8 цистернв виде уплощенных мешочков d = 0, 05–10 нм. Цистерны располагаются одна над другой наподобие стопки блинов на расстоянии 25–30 нм друг от друга, никаких структурных связей между ними нет. Число диктиосом варьирует от одной до нескольких тысяч в зависимости от вида клетки. Это образование имеет чашеобразную форму и тесно связано с гладкой ЭПС. Функции: 1) место сортировки, созревания, хранения, упаковки и выведения веществ, синтезированных в клетке; 2) образование секреторных гранул ( включений ); 3) посредник между ЭПС и цитоплазматической мембраной в процессе экзоцитоза;
4) участие в метаболических процессах ( синтез полисахаридов ) и химических преобразованиях ( синтез стероидов );
5) образование первичных лизосом; 6) участие в формировании клеточных мембран; 7) компартментализация.
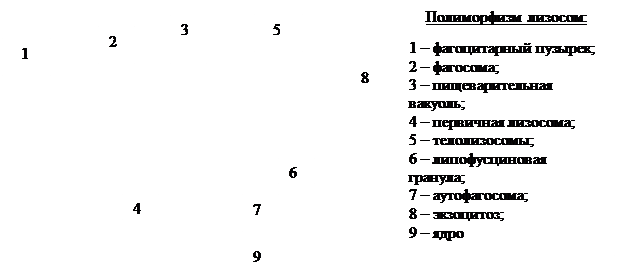
ЛИЗОСОМЫ Органоиды общего значения, имеющие одномембранный принцип строения. В 1949 году группа биохимиков во главе с Де Дювом открыла лизосомы как мембранные внутриклеточные частицы. Форма: сферическая. Размеры: 0, 2 – 1, 0 мкм. Структура: лизосома представляет собой пузырек, отграниченный от цитоплазмы однослойной мембраной, который несет более 60 протеолитических ферментов (20% из них сосредоточены на мембране лизосом, 80% – внутри).
Типы: 1) первичные лизосомы; 2) вторичные лизосомы ( пищеварительные вакуоли ); 3) остаточные тельца ( телолизосомы ); 4) аутофагосомы.
Первичные лизосомы – это пузырьки d=100 нм, отшнуровавшиеся от комплекса Гольджи и содержащие протеолитические ферменты (гидролазы). В дальнейшем, когда первичная лизосома сливается с фагосомой (пиносомой) – вакуолью, содержащей субстрат, она становится вторичной лизосомой. Внутри пищеварительной вакуоли происходит внутриклеточное переваривание. Судьба поглощенных биогенных веществ заключается в их расщеплении гидролазами до мономеров и в транспорте этих мономеров через мембрану лизосомы в состав гиалоплазмы, где они повторно утилизируются, включаются в различные синтетические и обменные процессы. Однако расщепление в ряде клеток может идти не до конца. Тогда в лизосомах накапливаются непереваренные продукты, в них нет ферментов, такие лизосомы получили название телолизосом. У человека при старении организма в клетках мозга, печени, мышечных волокнах в телолизосомах находят «пигмент старения» – липофусцин. Судьба остаточных телец может быть двоякой: одни выбрасываются из клетки путем экзоцитоза, другие (липофусциновые гранулы) остаются в клетках вплоть до их гибели.
Аутофагосомы по морфологии близки к вторичным лизосомам, но с тем отличием, что их субстратами являются фрагменты или даже целые цитоплазматические структуры (митохондрии, элементы ЭПС, рибосомы и др.).
Аутофагия – расщепление эндогенных биологических молекул, представляющих собой «сломанные», «изношенные» клеточные компоненты, что обеспечивает утилизацию в клетке баластных, дефектных органоидов или саморастворение органелл, особенно часто происходящее в условиях пищевого или кислородного голодания. Лизосомы имеют отношение к апоптозу (гибели клетки), когда клетка «переваривает» себя (аутолиз – саморастворение). Гетерофагия – расщепление экзогенных биологических молекул, попавших в клетку путем фагоцитоза (твердые вещества в упаковке) или пиноцитоза (капля в упаковке).
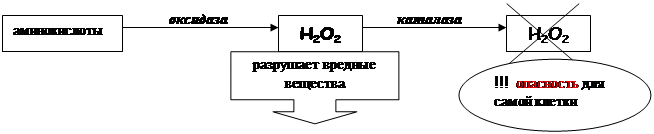
ПЕРОКСИСОМЫ (микротельца) Органоиды общего значения, имеющие одномембранный принцип строения. Впервые пероксисомы были выделены из печени и почек. Форма: сферическая. Размеры: у мелких 0, 15–0, 25 мкм, которые обнаруживаются во всех клетках; 0, 3–1, 5 мкм у крупных, которые находятся в клетках печени и почек. Структура: пероксисома – это округлое тельце, окруженное по периферии одним слоем мембраны. Внутри располагается сердцевина в виде кристаллов, представленных фибриллами или трубочками, и гранулярный матрикс. Во фракциях пероксисом обнаруживаются ферменты, связанные с метаболизмом перекиси водорода – это различные оксидазы, выделенные из сердцевины, и каталаза, содержащаяся в матриксе. Каталаза составляет до 40% всех белков в пероксисоме и играет важную защитную роль. Оксидазы катализируют образование пероксида водорода, который, будучи токсичным, разрушается затем под действием пероксидазы и каталазы. Эти реакции включены в различные метаболические циклы, например, обмен мочевой кислоты в клетках печени и почек.
Функция: 1) нейтрализация перекиси, которая является токсичным веществом для клетки; 2) депо для ряда ферментов, которые играют важную роль при превращении жиров в углеводы и катаболизме пуринов.
ВАКУОЛИ РАСТИТЕЛЬНЫХ КЛЕТОК Органоиды общего значения, имеющие одномембранный принцип строения и встречающиеся только в клетках растений. Популярное:
|
Последнее изменение этой страницы: 2016-08-24; Просмотров: 580; Нарушение авторского права страницы






 ②. ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ
②. ЖИДКОСТНО-МОЗАИЧНАЯ МОДЕЛЬ 






 Функция:
Функция: