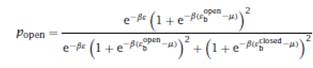|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Ионные каналы Revisited: лигандами каналов и MWC Модель
Существует много различных видов движущих сил, которые могут склонить чашу весов между закрытыми и открытыми состояниями. Лигандами каналы являются ионными каналами, у которых открытие и закрытие регулируется путем связывания лигандов с белком, который составляет канал. Одним из наиболее изученных примеров является никотиновая рецептора ацетилхолина, который играет роль в нервно-мышечном соединении. Она состоит из двух сайтов связывания ацетилхолина и равновесие между открытым и закрытым состоянием канала смещается в сторону открытого состояния путем связывания ацетилхолина. Изучить открытие канала в зависимости от концентрации ацетилхолина мы используем статистической механики модели канала, который является аналогом модели Моно-Wyman-Changeux (MWC) в dimoglobin, В случае dimoglobin, белок был в Т или R состоянии и лиганд имел более высокое сродство к двум сайтами связывания в состоянии R, которая сама по себе была выше по энергии, чем состояние T. Затем, путем изменения концентрации лигандов, можно было сдвинуть равновесие от Т до состояния R. Аналогичным образом, для лигандами канала, мы считаем, что открытое состояние канала, чтобы иметь более высокую энергию, чем закрытый канал, так что при отсутствии ацетилхолина канал закрыт. Имея ацетилхолин имеют более высокое сродство к каждому из двух участков связывания, когда канал находится в открытом состоянии, в отличие от закрытого состояния, мы еще раз имеем модель, в которой есть богатое взаимодействие между обязательными и конформационного состояния так же, как мы уже видели в случае гемоглобина. Для того, чтобы вычислить вероятность того, что канал будет открыт мы используем состояний и весов. Вероятность того, что канал открыт является суммой весов открытых состояний (WOPEN), деленное на сумму всех весов (WOPEN + wclosed), который производит формулу
Химический потенциал молекул ацетилхолина в растворе связанных с их концентрацией, eβ μ = (с / c0) eβ μ 0 и Поэтому мы можем переписать уравнение, как
Для дальнейшего упрощения формулы, мы ввели диссоциации константы Kopend= c0eβ (ε openb− μ 0) and Kclosed d= c0eβ (ε closedb− μ 0), для молекулы ацетилхолина связывания с каналом в открытых и закрытых говорится, соответственно. Мы можем использовать формулу для POPEN прокомментировать экспериментов, которые делают использование записей канала для измерения доли времени, что лиганд закрытого канала проводит в открытом состоянии в зависимости от концентрации ацетилхолина. Данные показывают две тенденции: открытие вероятность возрастает с увеличением концентрации ацетилхолина и концентрация, при которой вероятность достигает полувысоте различна для разных мутантных каналов. Наклоны в средней части графа, связанные со степенью кооперативности, также, кажется, показывают тенденцию к снижению. Модель MWC лиганд закрытого канала определяется тремя параметрами, разность энергий между открытым и закрытым состояниями, е, и двух констант диссоциации ацетилхолина, один для открытой и один для закрытого состояния рецептора. Если измерять концентрацию ацетилхолина в единицах Kclosed г, то есть ~c = с / Kclosed d, то есть два параметра слева, а именно, отношение двух диссоциации константы и коэффициент Больцмана, связанный с разностью энергий между закрытым и открытым состоянием канала. Это приводит к тому выражение для открытия вероятности вида
7.2. Скорость посттрансляционной модификации белков
7.3. Сколько белков синтезируется за одну трансляцию мРНК? ( 158 M)
Учитывая центральное место регуляции генов во всех областях биологии, существует большой интерес в определении переписи мРНК в клетках различных типов. Мы заинтересованы как в специфических генов и во всем транскриптома в зависимости от условий окружающей среды и стадии развития. Такие измерения обеспечивают прямое считывание мгновенного регуляторного состояния клетки в данный момент времени.
Суть оценки заключается в использовании признание того, что в течение клеточного цикла, количество белков должно быть удвоено за счет синтеза белка. Этот синтез белка на основе, в свою очередь, на распределение молекул мРНК, которые присутствуют в клетке. Как показано в этой задней части вычисления огибающей можно получить оценку для быстро делящихся клеток 103-104 мРНК в бактериальной клетке и 105-106 мРНК с ПЭО 3000 мкм3 характерным размером клетки млекопитающего. Один подход ориентирован на измерения " genomewide", в которой сделана попытка оценить и размеры количество мРНК по всей транскриптома. Эти результаты основаны на методе РНК-Seq, где отдельные мРНК упорядочиваются из клеточного лизата. Конечно, так как отсчет основан на частоте последовательности читает, она требует калибровки. Для достижения этой цели образец подсыпали стандартами мРНК, количество которой известно перед секвенированием. Результатом этого исследования состоит в том, что число транскриптов на клетку (≈ 8000 мРНК копий / клеточные) для клеток, выращенных в среде LB. примерно в три раза больше, чем количество транскриптов на клетку (≈ 3000 мРНК копий / клеток) для клеток выращенных в минимальной среде. Учитывая, что число генов превышает 4000, то это означает, что среднее число копий в LB составляет порядка одной на клетку. Для большинства генов это на самом деле еще меньше, Опять же, оба метода секвенирования и микроскопии были привлечены к нести на эти вопросы в дрожжах и других эукариот. Общее количество мРНК в клетке находится в диапазоне 20, 000-60, 000 в экспоненциально растущей бутонизации и делящихся дрожжей (BNID 104312, 102988, 103023, 106226, 106763). Как с нашими более ранними результатами для бактерий, и здесь мы видим, что каждый ген, как правило имеет только несколько молекул мРНК, присутствующей в клетке в любой момент времени. Для " типичных" клетках млекопитающих в кавычках значение 200000 мРНК в клетке (BNID 109916) в соответствии с нашей простой оценкой выше, и показывает, что масштабирование количество мРНК пропорционально размеру и скорости роста, как представляется, является полезным первое предположение.
Билет 8. 8.1. Случайные блуждания и структура макромолекул Chapter 8: Random Walks and the Structure of Macromolecules 311 Р
Есть много различных способов, характеризующих биологические структуры. Полезной альтернативой детерминистического описания структуры условия четко определенных атомных координат является использование статистической описания. Идея во всей науке, блуждание, и показать его полезность при характеристике биологические макромолекулы, такие как ДНК. Мы покажем, как эти идеи высшей точки в распределении вероятностей для расстояния от конца до конца полимеров и как это распределение может быть использовано для вычисления " Структура" ДНК в клетках, а также понимать одномолекулярной эксперименты, в которых молекулы ДНК (или белки) вытягиваются на и последующая деформация контролируется как функция от приложенного сила. Кроме того, мы покажем, как эти же идеи могут быть адаптированы думать о белках.
Популярное:
|
Последнее изменение этой страницы: 2017-03-08; Просмотров: 640; Нарушение авторского права страницы