|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Глава 5. ОБЩЕСТВЕННОЕ ПОВЕДЕНИЕ ЖИВОТНЫХСтр 1 из 12Следующая ⇒
Глава 5. ОБЩЕСТВЕННОЕ ПОВЕДЕНИЕ ЖИВОТНЫХ Общественное поведение — одна из важнейших форм видоспецифического поведения, изучение которого составляет обширный раздел этологии с момента ее возникновения и до настоящего времени. Благодаря многочисленным исследованиям типы организации сообществ (или социодемографических систем) и характер внутригрупповых отношений описаны достаточно подробно у большого числа видов. Наибольшее внимание при этом было уделено структуре сообществ у млекопитающих и птиц, однако мы рассмотрим также и такое замечательное явление, как общественное поведение насекомых (см.: 5.6). Исследование закономерностей существования и развития таких объединений у многих видов животных способствует пониманию этологической структуры популяций. Эти вопросы подробно проанализированы в монографии Е.Н.Панова (1983). Характер внутривидовых взаимоотношений особей тесно связан с различными типами их распределения в пространстве. Как правило, исследователи выделяют два основных типа использования территории. Первый — это не перекрывающиеся между собой и не защищаемые участки обитания отдельных особей или семей, второй — не перекрывающиеся, но защищаемые территории. Первый тип характерен для " социальных" видов животных, второй — для " территориальных" или одиночных видов (см.: 5.2.1). Даже у животных, относящихся к одиночным видам, существуют периоды жизненного цикла, когда они вступают в контакт с особями своего вида, например в сезон размножения. Взаимоотношения между самцами и самками, в особенности в брачный период, также относятся к категории внутривидовых общественных отношений. Минимальная социальная ячейка, цельность и самостоятельность которой поддерживаются в основном ее собственными, внутренними механизмами, называется микропопуляцией. Феномен микропопуляции особенно подробно изучался у домовой мыши, и применительно к этому виду микропопуляция называется " демом". По определению Н.П. Наумова (1977), подобное объединение особей именуется " парцеллой". Исследование динамики численности популяций показало важную роль поведения особей в этих процессах (Шилов, 1977). Видоспецифические особенности социальной организации более заметны в процессе формирования группы, а не в стабильный период ее существования, В качестве примера в разделе 5, 5.1 сравниваются социальные отношения и особенности агрессивных контшстов у нескольких близкородственных видов песчанок, Оригинальный научный подход, реализованный в этом исследовании (Гольцман и др., 1983; 1994), представляет собой, с одной стороны, прямое развитие классических этологических методов (наблюдение в естественной или " полуестественной" среде, сравнение близкородственных видов), а с другой — их принципиально новое развитое, поскольку он позволяет перенести внимание со статичной картины структуры группы на динамику ее формирования. Основные типы сообществ Сообществом называется такой тип внутрипопуляционных отношений, при котором особи образуют стабильные группировки, занимающие и защищающие определенную территорию. Они поддерживают постоянный обмен информацией, находятся в некоторых относительно постоянных отношениях и скрещиваются преимущественно друг с другом. Сообщество включает в себя, как правило, небольшое число особей. Это более мелкая, чем популяция, группа особей данного вида, которую иногда называют микропопуляцией. Для сравнительного изучения структурных и иных особенностей сообществ животных исследователям необходимо выделить признаки, по которым группировки особей разных видов могут различаться между собой. Важнейшими признаками при этом оказываются: 1) устойчивость группировок во времени; 2) взаимная координация действий особей в группе; 3) прочность связей между особями; 4) поддержание целостности группы (агрессия по отношению к " чужакам" своего вида). ОДИНОЧНЫЙ ОБРАЗ ЖИЗНИ Одиночно живущими называют животных, у которых контакты между особями поддерживаются преимущественно в период спаривания, а также (у некоторых видов) в период ухаживания за потомством. Наиболее характерными и достаточно известными примерами животных, живущих поодиночке, являются представители семейства кошачьих. Одиночный образ жизни можно считать устойчивой характеристикой большинства видов этой группы, за исключением львов. Следует отметить, что практически всегда он связан с поддержанием более или менее четко ограниченной территории, индивидуального участка, который служит местом пребывания особи и ее охотничьими угодьями. У многих крупных хищных млекопитающих (и птиц) размеры индивидуальных участков очень велики, поскольку они должны обеспечивать возобновляемые запасы дичи. Примером самого крупного одиночно живущего хищника средней полосы является бурый медведь (см.: 5.5.2). В социобиологических построениях (см.: 6.5.1) одиночный образ жизни рассматривается как исходный в эволюции социальных структур разной сложности (см.: Панов, 1983). Эволюцию усложнения социальной структуры, как правило, изучают на примере перепончатокрылых — группы насекомых, среди которых можно найти практически все переходы от одиночных к социально организованным формам. Примеры переходов от сугубо одиночного образа жизни к разным формам более или менее длительных семейных объединений у млекопитающих дают грызуны. У некоторых видов млекопитающих, например у волка, одиночный образ жизни составляет обязательную фазу годового жизненного цикла (см.: 5.5.2). АГРЕГАЦИИ, ИЛИ СКОПЛЕНИЯ Уже в ранний период развития этологии был обнаружен широкий диапазон различных типов социальной организации животных, начиная от одиночного образа жизни и кончая очень сложными сообществами приматов и общественных насекомых. Сообщества, даже самые примитивные, следует отличать от скоплений, или агрегаций, животных. Это такие объединения животных, которые формируются под действием какого-то физического фактора среды (пищи, температуры и т.п.). Примером агрегации могут служить стайки головастиков в прогретых солнцем местах водоема. Скопления характерны для многих видов беспозвоночных (см.: 5.6.1) АНОНИМНЫЕ СООБЩЕСТВА В книге " Агрессия", вышедшей впервые в 1966 г., К. Лоренц, рассматривая различные формы социальных структур, неявно проводит следующую классификацию: все сообщества животных можно разделить на два коренным образом различающиеся класса: анонимные, в которых нет ничего похожего на структуру (ни группировок, ни вожаков, ни ведомых), и персонифицированные, основанные на личных контактах, в которых возможно распределение ролей. Этот признак — наличие или отсутствие способности членов группы узнавать друг друга — используется в качестве одного из основных критериев классификации сообществ. Сообщество животных, персонально не знающих друг друга, называют анонимным. Было принято считать, что анонимные сообщества характерны главным образом для более низкоорганизованных групп животных, а с усложнением нервной системы и поведения в целом происходит и усложнение социальной организации. Однако это не совсем так. К таким анонимным сообществам относятся перелетные стаи, а также скопления многих видов птиц на ночевках, хотя последние могут состоять из более мелких групп, члены которых персонально знают друг друга. Следует отметить, что и в стае перелетных птиц (анонимном сообществе), например лебедей, диких гусей и журавлей, семейные группы (супружеские пары с детьми) держатся вместе и сохраняют личные связи. Но так бывает не у всех видов. В этой же ситуации аисты и цапли — члены супружеских пар — друг друга не узнают. Когда приходит время вить гнездо, супруги действуют независимо один от другого, даже если пара сохранилась: просто самец и самка каждый сам по себе прилетают на старое место. Анонимное сообщество часто демонстрирует удивительную сплоченность и целесообразность групповых действий. Так, стаи скворцов при появлении в воздухе ястреба-перепелятника или чеглока плотно стягиваются, спешат ему навстречу и, обтекая со всех сторон, вновь собираются у него в хвосте. Так же реагируют на хищника и многие рыбы. Хищники же, не только крупные, но и мелкие, не нападают на жертву внутри плотного стада, а стараются отбить кого-то одного или выждать, пока наиболее пугливая и беспокойная жертва сама отделится от стада (см.: Лоренц, 1994). В анонимных сообществах особи согласованно реагируют на биологически значимые сигналы, которые могут иметь разную модальность. В птичьих колониях это громкие крики, оповещающие о приближении хищника или просто о каких-то переменах обстановки. Сигналы другой модальности воспринимаются с помощью зрения, например беспокойные движения одной особи, которые побуждают остальных членов стаи следовать за ней. Такая реакция называется кинопсисом. Сигналы могут иметь химическую природу. Известно, например, что если в аквариум с рыбками налить немного воды из другого аквариума, в котором стайка таких же рыбок недавно спасалась от хищника, то подобная паника может охватить и обитателей первого аквариума, В данном случае информация передается с помощью особого вещества — феромона*) тревоги. *) Феромоны — это специфические вещества, выделяемые животными определенных половых и/или возрастных трупп и имеющие сигнальную функцию. Феромоны играют важную роль и в других сферах социального поведения (Новиков, 1985). В анонимном сообществе первыми обычно издают сигналы особи, оказавшиеся на периферии. Поскольку члены стаи все время меняют свое положение, то невольным лидером, за которым они устремятся, может оказаться любое животное, Такое сообщество называют эквипотенциальным. Анонимные сообщества, члены которых не проявляют агрессии по отношению к вновь присоединившимся особям своего вида, получили название открытых. К ним относятся, например, стада жирафов или кенгуру, которых привлекает вид сородичей, но они с равной легкостью присоединяются к данной группе, а затем покидают ее. Другой тип анонимного сообщества — закрытое. Его члены не различают друг друга " персонально", но могут выделять животных, принадлежащих к другому сообществу (" чужаков" ), например по запаху. К типичным закрытым анонимным сообществам относятся колонии крыс-пасюков (Rattus norvegicus). При появлении на территории колонии чужака все ее взрослые члены набрасываются на него и либо убивают, либо изгоняют. Единственный признак, по которому крысы отличают " своих" от " чужих", — это специфический для каждой колонии запах. Если крысу потереть подстилкой, взятой из другой колонии, она сейчас же будет убита сородичами, с которыми до этого жила в полном мире (рис. 5.2).
Рис. 5.2. Агрессивное столкновение между крысами-пасюками ИЕРАРХИЯ ДОМИНИРОВАНИЯ Структурированность сообщества — поддержание иерархической организации, обеспечивается прежде всего благодаря феномену доминирования и подчинения. Норвежский эколог Т. Шьелдерупп-Эббе (Schelderupp-Ebbe, 1924) обнаружил в группах домашних кур и уток строгую упорядоченность отношений между птицами. Каждая особь либо превосходит по силе партнера, либо уступает ему. Эти взаимоотношения были названы " порядком клевания". При формировании группы происходит " выяснение отношений" птиц друг с другом, в ходе которого постепенно выделяется одна, которая первой получает доступ к корму и гоняет от него всех остальных. Ниже ее на " лестнице доминирования" располагается птица второго ранга, которая превосходит всех, кроме главной, доминантной особи, и так далее. В самом основании находится особь, которую гоняют все члены группы. Такая иерархическая система вырабатывается при столкновении птиц в борьбе за " ограниченный ресурс" (за место на насесте, пищу), и на ранних этапах ее установления происходит много драк. Однако когда иерархия устанавливается, она оказывается стабильной, поскольку порядок соподчинения особей устойчиво поддерживается. Обычно при приближении высокоранговой птицы подчиненные особи уступают ей без сопротивления. Анализ показал, что предпосылкой стабильности иерархии является индивидуальное узнавание особей. В экспериментах, в которых одну и ту же птицу помещали последовательно в разные группы, " откалиброванные" таким образом, что она занимала в них разное иерархическое положение, куры обнаружили незаурядные способности к запоминанию и распознаванию членов каждой группы и без колебаний занимали причитающееся им положение. У кур иногда устанавливается совершенная линейная иерархия, так что ни одна птица никогда не клюет особей, стоящих выше ее на иерархической лестнице (табл. 5.1). Подобные " идеальные" сообщества исключительно редки. Среди беспозвоночных их образуют, например, сверчки и речные раки, у которых иерархические отношения также строятся на основе индивидуального распознавания. В то же время у большинства видов животных обнаруживаются различные отклонения от строгого линейного порядка. Описанное явление — формирование иерархической структуры группы — стало рассматриваться как механизм, благодаря которому одно или несколько животных получают приоритет во всех жизненных ситуациях. Предполагалось, что иерархическое ранжирование выделяет наиболее жизнеспособных особей, обеспечивая преимущественный успех их потомства в процессе естественного отбора. Действительно, некоторые эксперименты прямо свидетельствовали о лучшей приспособленности членов жестко организованных сообществ. Например, в одних группах кур доминантную особь регулярно удаляли и заменяли незнакомой птицей, так что остальные вынуждены были постоянно заниматься " выяснением отношений", тогда как членов контрольных групп не тревожили. В контрольных группах агрессивные стычки происходили реже, яйценоскость кур была выше, т.е. группы с постоянным составом обладали явным преимуществом благодаря стабильной социальной среде. Аналогичную иерархическую структуру и свойства имеют колонии серых крыс (см.: 5.5.1).
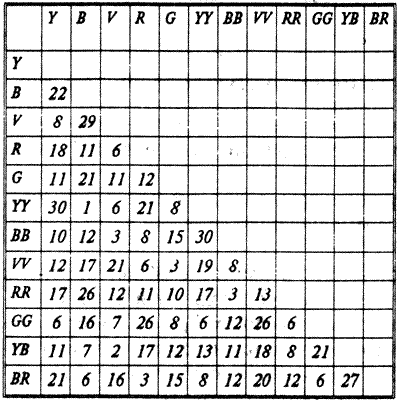
Таблица 5.1 " Идеальная" линия иерархии в группе, состоящей из 12 кур Один из наиболее распространенных методических приемов, в особенности при исследовании физиологических основ доминирования и агрессивности, — это экспериментальный анализ поведения в " конкурентных ситуациях", когда моделируется ограниченность доступа к ресурсам. Для этого двум животным, на определенное время лишенным пищи или воды, одновременно открывают путь к одной кормушке или поилке. В зависимости от условий эксперимента подкрепление может получить либо только один из конкурентов, либо доминантом считается животное, которое больше съедает или дольше контролирует кормушку. Эксперименты такого рода, проведенные на животных разных видов, показали, что корреляция между разными показателями доминирования (конкуренция за пищу, воду, территорию, доступ к половому партнеру, возможность уйти в гнездо и т.п.) может быть слабой или совсем отсутствовать. Даже в строго контролируемых лабораторных условиях при использовании генетически однородных однополых и одновозрастных животных не найдено " единого" доминанта. РИТУАЛЫ И ДЕМОНСТРАЦИИ Поддержание структуры сообщества осуществляется на основе агонистического поведения, т.е. комплекса реакций нападения, угрозы, подчинения и бегства. Как уже упоминалось, установление иерархии может происходить в результате прямых столкновений и драк, тогда как по мере ее упрочения в большинстве случаев прямые проявления агрессии уступают место угрозам и демонстрациям. Выше (раздел 4.3) были рассмотрены типичные примеры угрожающих демонстраций у рыб и чаек. Согласно гипотезе Н.Тинбергена, выраженность практически всех демонстраций определяется соотношением уровня агрессивности и страха, которые в данный момент переживает животное (рис. 5.5 и 5.11).
Рис.5.5. Позы угрозы у обыкновенной чайки, связанные с разным соотношением тенденций к атаке (темные столбцы) и к избеганию (светлые столбцы) (по Мак-Фарленду, 1988} Знаменитая серия зарисовок немецкого этолога П.Лейхаузена показывает мимику домашней кошки при разных сочетаниях уровней агрессии и страха (рис. 5.6): агрессивная тенденция усиливается слева направо, страха — сверху вниз.
Рис. 5.6. Изменения мимики кошки в соответствии с разными уровнями тенденции к агрессии и реакции страха (по П. Лейхаузену) Другую группу демонстраций составляют ритуализованные движения, тормозящие агрессию со стороны более сильных сородичей. Их называют позами " покорности" или " умиротворения". По всем своим параметрам они противоположны жестам угрозы. Так, коралловая рыбка, угрожающая сородичу, демонстрирует ему свой яркий наряд, распахивает плавники или жаберные крышки, движется резко, демонстрируя силу. Когда же она " просит пощады", все выглядит совершенно по-другому; ее окраска бледнеет, она прижимает плавники, поворачивается к сопернику узким сечением тела, стараясь казаться маленькой, движется медленно, крадучись. Подобным же образом, демонстрируя угрозу, птицы распушают перья, чтобы казаться как можно крупнее, тогда как в знак подчинения они прижимают перо, и их силует становится более тонким. У макаков-резусов позы и движется доминантного и подчиненного животных диаметрально различны (рис. 5.7 и 5.11).
Рис.5.7. " Стиль" поведения доминантных и подчиненных особей макаков-резусов (Меннинг, 1982) Выразительные позы угрозы и подчинения у воронов (рис. 5.8) не оставляют сомнений в их функциональном " назначении".
Рис. 5.8. Ворон в позе угрозы и в позе подчинения (по Хейнриху, 1995)
Рис.5.9. Позы подчинения у волков. Формы ритуалов умиротворения многообразны, тем не менее некоторые из них (поза подставления, реакция выпрашивания корма) у самых разных групп животных одинаковы (рис. 5.9). К. Лоренц (1994) выделил несколько основных особенностей таких ритуалов: 1. Демонстративное подставление наиболее уязвимой части тела. Так, побежденный волк отворачивает голову и подставляет победителю чрезвычайно ранимую боковую сторону шеи, выгнутую навстречу укусу. Галка подставляет под клюв птицы, которую нужно умиротворить, свой незащищенный затылок — обычную мишень при серьезном нападении с целью убийства. У многих видов птиц оперение на таких участках тела имеет особую окраску. У галок оно шелковисто-серое, а у тех врановых, которые крупнее и темнее на затылке имеется более светлое пятно. По данным Н.Тинбергена (1974), чайки, демонстрируя умиротворение, отворачивают от противника клюв (см.: рис. 4.6 в), при этом привлекает внимание их белоснежный затылок (как у обыкновенной чайки) или особый рисунок на светлом фоне (как у молодой моевки). 2. Воспроизведение некоторых элементов детского поведения. Распространенный вариант ритуала умиротворения у птиц — это имитация позы птенца, выпрашивающего корм (рис. 5.10).
Рис 5.10. Ритуал " выпрашивания корма" у воронов а — рисунок (по Хейнриху, 1995), б — фотография Хейнриха В репертуаре социального поведения псовых жесты умиротворения также играют важную роль; они происходят непосредственно из отношений щенка с его матерью и являются производными от движений сосания или просьбы накормить (см.: рис. 5.9). Как предполагает К. Лоренц (1994), эти действия превратились в ритуалы у тех животных, у которых изначально существовали более общий, социальный смысл, оказалось возможным формирование крупных социальных групп на базе супружеских пар и семей.
Рис.5.11. Сигнальные позы у клуши 3. Выражение социальной покорности с помощью действий, характерных для поведения самки при спаривании. Эти действия типичны для многих обезьян, особенно павианов. Ритуальный поворот задней части тела означает признание более высокого ранга другой обезьяны. Позу подставления они выполняют с самого раннего возраста без всякого обучения, даже при воспитании в изоляции от сородичей (рис. 5.12). Как уже упоминалось, те части тела, которые животные демонстрируют в знак угрозы или подчинения (релизеры — см.: 3.4), окрашены особенно ярко и заметно, чем подчеркивается выразительность церемонии. Так, у некоторых видов павианов задняя часть тела, которую они демонстрируют противнику в позе подставления, раскрашена самым фантастическим образом.
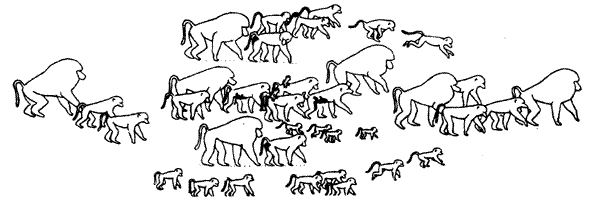
Рис 5.12 Поза " подставления у самца павиана занимающего " подчиненное" положение СЛОЖНЫЕ СИСТЕМЫ ИЕРАРХИИ Представление о том, что в условиях ограниченного доступа к какому-либо ресурсу доминирование способствует его получению в первую очередь, подтверждено в многочисленных исследованиях социального поведения животных (подробнее см.: Резникова, 1997). С ростом числа исследований на животных разных видов выявилось множество отклонений от классической линейной схемы иерархической структуры сообществ. Наиболее распространенное отклонение — превращение " лестницы" или " цепочки" в двухуровневую структуру: доминант — и все остальные. Случаи такого абсолютного доминирования наблюдаются, например, при содержании самцов домовых мышей или самцов колюшки в ограниченном пространстве. В иных случаях в иерархических " цепочках" могут наблюдаться " петли" или " треугольники", когда особь А гоняет особь В, особь В — особь С, но А боится С. Описаны даже такие ситуации, когда длинная иерархическая цепочка замыкалась в круг. Показано, что в сообществах могут действовать параллельные системы иерархии — у самцов и у самок. Изменение ранга самки, как это было описано К. Лоренцем (1978) при изучении сообщества галок, может происходить, если она становится супругой высокорангового самца. У большинства видов самцы имеют более высокие ранги, однако имеются и исключения. У некоторых видов животных внутри большого сообщества существуют стабильные группировки — кланы, чаще всего образованные одной или несколькими самками с их потомством одного-двух поколений (см. Крук, 1977) Такие " материнские кланы" можно наблюдать, например, у слонов (И. и О.Дуглас-Гамильтон, 1981), дельфинов (Прайер, 1981, см. также обзор. Крушинская, Лисицына, 1983), гиеновых собак (Дж. и Г. Лавик-Гудолл, 1977). На рис. 5.13 приведена схема " построения" группы павианов при перемещении на открытом пространстве.
Рис 5.13 Стая павианов на марше через открытую равнину Помимо относительно стабильных группировок и кланов, члены сообщества могут образовывать и временные коалиции, направленные либо на достижение " сиюминутной выгоды", либо на то, чтобы повысить свой общий ранг. Множество примеров подобных ситуаций дают исследования приматов (японских макаков, зеленых мартышек, павианов, шимпанзе). Так, два—три самца могут объединиться, чтобы отвлечь доминанта, инсценируя драку или интересную находку, и, пока доминант " наводит порядок", по очереди получить доступ к пище или даже увести самку. Пример такой же " эксплуатации" молодым самцом принятого в сообществе доброжелательного отношения ко всем детенышам, можно видеть на рис. 5.14.
Рис. 5.14. Неполовозрелый самец варварийской обезьяны (берберийского макака) и детеныш этого вида. Сначала они сидят недалеко друг от друга (вверху), Затем самец подманивает детеныша, делает ему дружественное гримасу, детеныш к нему подходит (в середине), забирается к самцу на спину, и тот может отнести и " предъявить" его старшему самцу. Благодаря этому он может остаться рядом с доминантом в относительной безопасности Существует достаточно много примеров, когда в сообществе приматов складывается не столько иерархия особей, сколько иерархия группировок (Гудолл, 1992). КАК РАБОТАЮТ СИГНАЛЫ Известно, что разные группы животных более или менее специализированы по сенсорной модальности используемых сигналов, в зависимости от степени развития у них тех или иных органов чувств. Так, тактильная коммуникация доминирует во взаимодействиях многих беспозвоночных, например у слепых рабочих термитов, которые никогда не покидают своих подземных туннелей, или у дождевых червей, которые ночью выползают из нор для спаривания. У беспозвоночных осязание тесно связано с химической чувствительностью, так как специализированные тактильные органы, например антенны насекомых, часто снабжены хеморецепторами. Общественные насекомые передают большое количество информации путем комбинации тактильных и химических сигналов. Тактильная коммуникация в силу своей природы возможна только на близком расстоянии. Длинные антенны тараканов и раков позволяют им исследовать мир в радиусе одной длины тела, но это почти предел для осязания. Тактильная коммуникация сохраняет свое значение у многих позвоночных, в частности у млекопитающих, наиболее " общественные" виды которых проводят значительную часть времени в физическом контакте друг с другом. У приматов важнейшую форму социальных контактов составляет обыскивание шерсти, или " груминг". Другие сенсорные системы — системы зрения, слуха и обоняния обеспечивают общение на значительном расстоянии. Звук и запах имеют дополнительное преимущество благодаря тому, что они способны преодолевать естественные препятствия, например густую растительность. Звуковые сигналы. Сигналами, рассчитанными на большое расстояние, являются обычно крики. Например, территориальные птицы поют свои песни, выбирая самую высокую точку местности (" песенный пост" ). Птицы открытых ландшафтов (жаворонки, луговые коньки) поют, летая высоко над своей территорией. Рекорд дальности в звуковой коммуникации животных поставлен горбатыми китами: их песни могут восприниматься другими китами, находящимися на расстоянии нескольких десятков километров. Химические сигналы особенно хорошо развиты у насекомых и у млекопитающих. Феромоны бабочек улавливаются самцами с подветренной стороны за 4—5 км, и они самые устойчивые из феромонов насекомых. Наоборот, феромоны тревоги, используемые муравьями, распространяются на расстояние в несколько сантиметров и быстро распадаются, что вполне объяснимо, так как эти сигналы призваны обеспечить быструю, но локальную реакцию. Многие млекопитающие маркируют свою территорию при помощи запаховых меток, часто концентрируя их на особых местах. Запах используется также для сообщений о состоянии самок млекопитающих в период размножения. Во время эструса они выделяют специальные химические вещества, сигнализируя этим готовность к встрече с самцом. Этот тип сигнала используется у самых разных видов. Он аналогичен, например, запахам, издаваемым неоплодотворенными самками бабочек и привлекающим самцов с весьма далеких расстояний. Такие вещества называются половыми феромонами (см. также: 5.2.3). Зрительные сигналы могут действовать лишь на относительно коротком, в пределах видимости, расстоянии. Исключение составляют простые сигналы тревоги, в виде белых пятен на теле, например хвосты оленей и кроликов, видимые на большом расстоянии. К зрительным сигналам относятся также широко распространенные опознавательные знаки, многие из которых действуют по принципу " есть или нет". Так, ключевым признаком для определения пола у американского золотистого дятла является наличие черных усиков у самца, и если вклеить их самке, как это сделал Г.Нобль, самец сразу прогоняет ее (рис. 5.17). Ритуальные демонстрации и танцы брачных партнеров также воздействуют в первую очередь на зрительный анализатор (рис. 5.18).
Рис. 5.17. Самец (слева) и самка (справа) золотистого дятла. Ключевой признак для определения пола птицами своего вида — черные усики самца
Рис. 5. 18. Совместная брачная демонстрация у манакина. Часть демонстрации заключается в " кувыркании" двух самцов перед самкой (по Alcock, 1989) О. Меннинг отмечает связь частоты использования цветовых зрительных сигналов с развитием цветного зрения у животных. Сложная брачная демонстрация бриллиантовой амадины включает в себя и " преподнесение подарка" (рис. 5.19). Оно практически универсально для многих групп, за исключением большинства млекопитающих. В ранней истории млекопитающих, когда их предки были ночными насекомоядными, цветовое зрение давало им мало преимуществ и почти утратило роль в процессе коммуникации. Поэтому яркая разноцветная раскраска некоторых рыб, рептилий и птиц поразительно контрастирует с универсальной серой, черной и коричневой окраской млекопитающих. Исключение составляют некоторые белки и приматы — это как раз те две группы млекопитающих, у которых цветовое зрение более развито, чем у остальных. В их окраске встречаются синие, красные, зеленые и желтые участки, которые они могут демонстрировать, например, в ритуалах угроз или подчинения.
Рис.5.19. Ухаживание у бриллиантовой амадины Steganopleura guttata. Самец демонстрирует самке подарок — травинку (а), поет и выполняет танец (б), а затем, сохраняя травинку в клюве, принимает позу выпрашивания пищи, характерную для молодых птиц (в) (по Eibl-Eibesfeldt, 1977) В естественных условиях сигналы различных модальностей часто объединены в эффективные комбинации, сочетающие, например, и звук, и зрительный стимул. Хорошим примером являются брачные ритуалы райских птиц, включающие характерные позы, демонстрации " ритуальных" перьев, прыжки, крики и хлопанье крыльями. Один из примеров такой комбинации, также действующей в сфере полового поведения, — строительство и украшение шалашей, которые служат местом интенсивного соперничества между самцами у 14 из 18 видов шалашников, или беседковых птиц (сем. Ptilonorhynchidae), населяющих Австралию и Новую Гвинею. У двух видов — это беседка, достигающая полутораметровой высоты, в центре которой находится разукрашенное птицей деревце, а сводчатый коридор ведет из беседки на расчищенную площадку для тока. Для украшения беседки используются перья, раковины, кусочки костей, надкрылья жуков и цикад, а возле поселений человека еще и монеты, булавки, стекло, бижутерия, чайные ложечки, гвоздики, винтики, бумага и тому подобные предметы, сочетающиеся по цвету с украшениями природного происхождения (рис. 5.20). У некоторых видов самцы к тому же раскрашивают шалаши, смешивая сок ягод со слюной. Это пример, по-видимому, одного из самых сложных зрительных сигналов (см. также: Резникова, 1997).
Рис. 5.20. Самец австралийского атласного шалашника у своего шалаша (рис. О. Перепелкиной) Таким образом, сигналы, используемые животными, весьма многочисленны. Однако все их многообразие у разных видов укладывается приблизительно в 10 основных категорий: До сих пор мы рассматривали работу различных сигналов, не обсуждая вопроса о наличии четких границ, разделяющих отдельные единицы коммуникации. Ведут ли животные диалог по принципу " вопрос-ответ", или сигналы плавно переходят один в другой, образуя континуум, в котором партнеры могут выделять отдельные фрагменты и реагировать на них? Этологи высказывали представление о том, что одни сигналы могут действовать как триггеры (" спусковые крючки" ), а другие — как " насосы". Крики тревоги, например, постоянны для каждого вида. Они действуют как триггеры, резко меняя поведение особи. О принципе " насоса" говорят тогда, когда влияния сигналов постепенно накапливаются и изменяют вероятность того, что получатель будет отвечать. Например, самцы голубей при ухаживании в течение нескольких дней много раз повторяют характерные " поклоны", прежде чем произведут на самку нужное впечатление. В этом случае градуированное воздействие заключается в накоплении эффекта от повторения. Наряду с этим есть сигналы, которые сами по себе могут быть градуированными. Так, при агрессивных стычках животные нередко испускают различные крики или осуществляют зрительные демонстрации (движения хвоста у собак, приподнимание хохолка у соек). Эти сигналы имеют несколько градаций, которые коррелируют с вероятностью того, что животное нападет или убежит. Приведенный выше рис. 5.6, на котором показаны соответствующие " градации" в мимике кошки (по Лейхаузену), представляет собой иллюстрацию этого положения. Популярное:
|
Последнее изменение этой страницы: 2017-03-11; Просмотров: 2306; Нарушение авторского права страницы