|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Метаболизм микробной клетки и его влияние на биотехнологию производства лекарственных средств
Известно, что рост клетки, ее нормальное функционирование и даже само выживание зависит от того, насколько эта клетка обладает способностью управлять процессами биосинтеза, а также вносить качественные преобразования в работу метаболического аппарата, отвечая на изменение условий среды. Такие функции клетки потребовали развития и наследственного закрепления весьма сложных и тонких регуляторных механизмов, обеспечивающих прежде всего экономичность метаболических процессов и, конечно, высокий уровень их координации. Часто цели биотехнологов, направленные на усиление образования того или иного продукта, или синтез нового продукта, естественно встречают сопротивление клетки, желающей сохранить свою стабильность, что влечет за собой изменение регуляторных механизмов клетки. В микробной клетке в процессе ее роста и жизнедеятельности происходит огромное число реакций, катализируемых ферментами. Участниками таких реакций могут быть первичные и вторичные метаболиты. К первичным метаболитам относятся аминокислоты, сахара, аминосахара, структурные белки, липиды, пурины, пиримидины и др. Вторичные метаболиты представляют вещества, образующиеся в ответ на стрессовую ситуацию, изменение экологии, например, это антибиотики, феромоны.
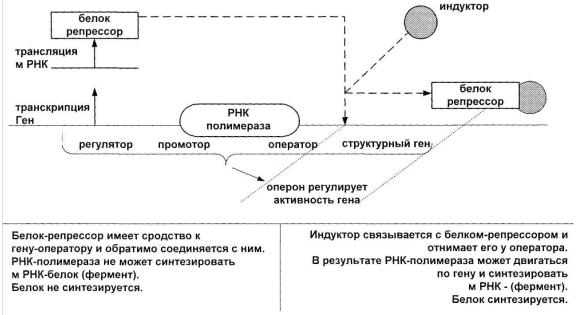
Индукция и репрессия синтеза ферментов. Механизмы индукции и репрессии предохраняют клетку от напрасной траты аминокислот и энергии на образование ненужных в данных условиях ферментов. С другой стороны, если появляется необходимость, эти ферменты могут быстро синтезироваться. Из многих тысяч ферментов микробные клетки в процессе роста способны синтезировать постоянно и не зависимо от питательной среды так называемые конститутивные ферменты (примером таких ферментов являются ферменты гликолиза, превращающие глюкозу в пируват). Другие ферменты, называемые адаптивными или индуцибельными, образуются только тогда, когда их субстраты (или структурные аналоги субстратов) присутствуют в среде. Например, клетки Е.соli., которые растут на среде с глюкозой, содержат только следы ферментов метаболизма лактозы. Если же эти клетки перенести в среду, содержащую в качестве единственного источника углерода лактозу, то можно наблюдать значительное повышение активности бетагалактозидазы. Этот фермент способен гидролизовать лактозу на D-галактозу и D-глюкозу. Это пример индукции фермента. Клетка получает возможность полностью усвоить лактозу в результате последовательной индукции ферментов, которые превращают лактозу в метаболиты, непосредственно используемые клеткой. Индукция фермента – это относительное увеличение скорости синтеза фермента в ответ на появление химического соединения – индуктора. Схема индукции Жакобо и Моно. Явление индукции ферментов впервые было изучено лауреатами Нобелевской премии Ф. Жакобом и Ж.. Моно.
Рис.7. Схема индукции фермента
В 1961 году Ф.Жакоб и Ф.Моно на основании генетического и биохимического изучения усвоения лактозы клетками Е.соli. предложили гипотезу о регуляции активности генов у бактерий, получившую широкую известность как «модель оперона». Согласно этой модели, на хромосоме имеется по крайней мере четыре компонента системы регуляции: структурный ген (или гены, контролирующие связанные между собой биохимические функции), ген-регулятор, оператор и промотор, которые составляют оперон. Ген-регулятор (R), определяет структуру белка-репрессора. Этот белок способен связываться с оператором (О), который контролирует функционирование расположенных рядом структурных генов (S 1, S 2, S 3). Промотор (Р) является начальным участком для связывания РНК-полимеразы, представляющей фермент, который катализирует транскрипцию ДНК в мРНК. Если белок-репрессор связан с оператором, то РНК-полимераза не имеет возможности перемещаться (или присоединяться) к промотору и в этом случае мРНК, которые комплементарны последовательности генов S 1, S 2, S 3, не образуются. Следовательно, и соответствующие ферменты также не синтезируются. В другом случае, когда оператор свободен от белка-репрессора, РНК- полимераза, присоединившись к промотору (Р), может перемещаться и транскрибировать гены S 1, S 2, S 3. Образование индуцибельных ферментов происходит при добавлении индуктора, который связывается с белком-репрессором и инактивирует его. Жакоб и Моно считали, что репрессор представляет собой аллостерический белок, который содержит два специфических центра. Один центр обладает сродством к нуклеотидной последовательности оператора, а другой центр обладает сродством к молекуле индуктора. Присоединение индуктора к репрессору ведет к снижению сродства первого аллостерического центра к оператору, в результате оператор освобождается от репрессора. Мутации в гене-регуляторе или в операторе могут нарушать образование репрессора или его связывание. В обоих случаях потребность в индукторе для синтеза ферментов исчезает. Такие мутанты называются конститутивными, так как у них соответствующие ферменты синтезируются постоянно. Получение конститутивных мутантов имеет важное значение в селекции промышленных штаммов микроорганизмов. Аллостерический центр (определение, функционирование). Исследование механизма подавления под действием конечного продукта, проведенные in vitro с использованием очищенных ферментов показали, что ингибитор образует комплекс с ферментом, связываясь со специфическим участком, полностью отличающимся от активного центра фермента. Этот участок фермента получил название аллостерический участок или аллостерический центр (от греческого «аллос»-другой, «стереос» - пространственный), а ферменты, имеющие аллостерический центр, - аллостерическими ферментами. Аллостерический центр – это участок фермента, который образует комплекс с конечным продуктом, в результате чего искажается трехмерная структура фермента (в том числе и его активного центра) и фермент становится не способным катализировать реакцию. Аллостерические ферменты представляют собой олигомеры, состоящие из двух, четырех, шести (или более) идентичных или различных субъединиц, способных взаимодействовать друг с другом. Связывание ингибитора искажает трехмерную структуру фермента. Это искажение передается активному центру и вызывает подавление активности фермента. Таким образом, некоторые метаболиты обладают способностью передавать информацию (как правило, путем изменения концентрации) ключевым ферментам о состоянии обмена веществ в клетке, в частности, подавать сигнал о необходимости прекращения дальнейшего функционирования данного метаболического пути. При мутационном повреждении аллостерического центра процесс биосинтеза не будет более подавляться конечным продуктом, и этот продукт начнет выделяться в среду. Для отбора таких мктантов используют структурные аналоги метаболитов. Например, 5-метилтриптофан, аналог триптофана, так же как и триптофан, подавляет антранилатсинтетазу, но не заменяет триптофан в белке и поэтому задерживает рост бактерий. Для общего обозначения регуляторных молекул, которые ингибируют или активируют аллостерические ферменты, используют термин аллостерические эффекторы. Ретроингибирование (схема, меры борьбы). Получение мутантов, устойчивых к аналогам метаболитов, часто используют в селекции продуцентов аминокислот, нуклеотидов и витаминов. Активные и неактивные формы фермента могут различаться также наличием или отсутствием каких-либо химических групп, ковалентно связанных с белком. Взаимный переход фермента из одной формы в другую достигается путем фосфорилирования –дефосфолирирования (ферменты метаболизма гликогена эукариотических клеток), аденилирования – деаденилирования (глутаминсинтетаза Е.соli), ацетилирования –деацетилирования. Ковалентную модификацию можно рассматривать как частный случай аллостерической регуляции. Наиболее гибким и широко распространенным способом контроля метаболизма в клетке является регуляция активности фермента по принципу обратной связи. Известно, что процессы биосинтеза многих незаменимых (первичных метаболитов) характеризуются тем, что конечный продукт данного (конкретного) биосинтетического пути при повышении его концентрации подавляет активность первого фермента этого пути. В результате такого подавления и соответствующий процесс биосинтеза останавливается. Конечный продукт, а также и промежуточные продукты, участвующие в его образовании, не накапливаются в клетке. Этот механизм автоматической регуляции называют подавлением под действием конечного продукта или ретроингибированием.
Рис. 2. Ретроингибирование по принципу обратной свяязи
Это высокоспецифическое подавление активности первого фермента заключительного этапа пути биосинтеза триптофана обеспечивает строгую и очень гибкую регуляцию новообразования этой аминокислоты в зависимости от скорости включения ее в белок и присутствия в ростовой фазе. В процессе роста бактерий преимущественно используют добавление аминокислот, пуринов, пиримидинов, так как эти соединения оказывают ингибирующее действие на свой собственный синтез из молекул предшественников. Борьба с ретроингибированием 1. Создание мутантных штаммов, которых фермент бы не имел аллостерического центра. 2. Удаление накапливающегося целевого продукта: ферментацию в этом случае осуществляют с сорбентом (активированным углем), на котором триптофан сорбируется. 3. Действие на аллостерический центр специальных активатором – аллостерических эффекторов, которые более активно связываются с аллостерическим центром, чем ингибитор (триптофан).
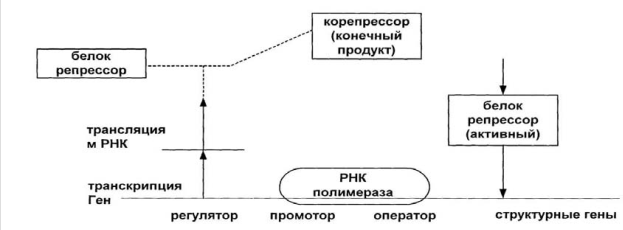
Катаболитная репрессия Катаболитная репрессия – это подавление конечным продуктом образования ферментов, катализирующих реакции образования конечного продукта.
Рис.8. Схематическое изображение катаболитной репрессии
В природных условиях в среде может находиться одновременно несколько субстратов, которые микробная клетка способна усваивать в качестве источников углерода и энергии. Однако это не приводит к синтезу всех ферментов, необходимых для катаболизма. В первую очередь образуются те ферменты, которые обеспечивают утилизацию наилучшего субстрата для поддержания наиболее высокой скорости роста. Для многих микроорганизмов таким субстратом является глюкоза. Клетка усваивает ее в первую очередь. Таким образом, глюкоза может влиять на утилизацию других субстратов, вызывая у микроорганизмов катаболитную репрессию. Глюкоза относится к легко усвояемым или быстро ассимилируемым субстратам, которые вызывают постоянную более или менее выраженную репрессию катаболитических ферментов. В результате подавляется окисление других субстратов. Катаболиты глюкозы (АТФ) накапливаются внутри клетки и подавляют синтез катаболитических ферментов. Помимо репрессии имеет место и процесс ретроингибирования – подавление активности ключевого фермента по принципу обратной связи – АТФ подавляет активность фермента гликолиза фосфофруктолазы, связываясь с ее аллостерическим центром. • Для биотехнолога важно иметь такие мутанты, у которых не было бы процесса катаболитной репрессии, например, она может неблагоприятно отразиться на синтезе антибиотиков. • Генные инженеры при получении новых продуцентов должны обращать внимание на механизм репрессии. Если репрессия синтеза конечного продукта имеет место, то можно получить штаммы, у которых этот механизм нарушен. У таких штаммов ген-оператор не реагирует с репрессором. |
Последнее изменение этой страницы: 2017-04-12; Просмотров: 761; Нарушение авторского права страницы