
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Глава 4 ФОРМЫ ИНДИВИДУАЛЬНОГО ПОВЕДЕНИЯСтр 1 из 11Следующая ⇒
Глава 4 ФОРМЫ ИНДИВИДУАЛЬНОГО ПОВЕДЕНИЯ В этой главе мы рассмотрим некоторые формы поведения, проявляемые отдельными особями в процессе адаптации к окружающей их среде. Это будут действия, с помощью которых животное получает пищу, воду, кислород и другие источники энергии; поддерживает чистоту своего тела; взаимодействует с окружающей средой, а также определенным образом организует свою активность. Все эти формы поведения имеют решающее значение для выживания. Они связаны с рядом конкретных функций, к которым относятся: 1) локомоция; 2) питание и дыхание; 3) терморегуляция; 4) поиски убежища; 5) избегание хищников; ч 6) сон; 7) поддержание чистоты тела; 8) выделение; 9) исследовательская активность; 10) игра; 11) использование орудий; 12) биологические ритмы.
Репродуктивному и общественному поведению будут посвящены главы 5 и 6. Здесь нужно сделать три замечания. Во-первых, при распределении материала по трем главам этой части и по разделам внутри глав мы исходили из соображений удобства и ясности изложения. Многие разграничения несколько произвольны, а некоторые из них в лучшем случае неоднозначны. Например, поиски укрытия могут иметь важное значение для того, чтобы избежать хищников; исследовательское поведение может составлять часть игры; потребление пищи влияет на терморегуляцию. Практически все эти типы поведения (например, питание, поиски укрытия, сон) иногда могут носить ярко выраженный общественный характер. Даже акты выделения могут иметь большое социальное значение, как, скажем, использование мочи для того, чтобы пометить занятую территорию. Таким образом, функциональное подразделение проводится здесь исключительно для удобства изложения, и его не следует принимать как безусловное. Во-вторых, следует помнить, что «животное – это приспособленный организм, а не просто совокупность специализированных адаптации» (Bunnell, 1973, р. 79). Способы приспособления животного к различным аспектам окружающей его среды переплетаются и взаимодействуют, образуя единое функциональное целое. Например, способ питания данного организма влияет на характер передвижения, на способ избегания врагов и на особенности сна и в свою очередь подвержен влиянию каждого из этих видов поведения. Решающий критерий – выживание, а размножение служит «нижним пределом». Действие отбора направлено на создание целостных организмов, способных функционировать в среде, где они обитают, а не на полное совершенство в какой-либо отдельной области. В-третьих, совершенно очевидно, что исчерпывающим образом изложить все эти вещи в одной главе невозможно. Мы попытаемся лишь дать некоторое представление о разнообразии форм поведения в пределах данной его категории, а также приведем по нескольку интересных примеров. Локомоция Локомоция имеет большое значение для большинства видов поведения, . так как она позволяет животному перемещаться в пространстве. Типы локомоции Существует пять основных типов локомоции: 1) движение при помощи жгутиков или ресничек; 2) амебоидное движение, осуществляемое путем изменения формы тела; 3) волнообразное движение; 4) реактивное движение и 5) движение при помощи конечностей (Maier, 1970). При движении первого типа животное передвигается в результате биения либо одного жгутика, либо группы волосовидных ресничек; такое движение широко распространено у простейших У беспозвоночных часто встречается также перемещение путем изменения формы тела, например путем вытягивания псевдоподий у амебы. Движение за счет волнообразных сокращений, пробегающих по телу, особенно характерно для змей, водных млекопитающих и рыб. При реактивном движении вода с силой выталкивается из организма в окружающую среду; этот тип локомоции используется у ряда беспозвоночных, таких, как медузы и кальмары Передвижение с помощью конечностей, будь то ноги, крылья или плавники, характерно для большинства позвоночных и для некоторых беспозвоночных. Передвижение при помощи конечностей Локомоция в результате движения конечностей может осуществляться в воде, на деревьях, в воздухе, под землей или на поверхности земли В воде. Плавание с использованием конечностей свойственно моржам и многим другим позвоночным. Разнообразные млекопитающие, живущие в основном на суше, в том числе многие грызуны, обезьяны и хищники, в случае необходимости достаточно хорошо плавают. На деревьях. Для большинства видов, обитающих на деревьях, характерно наличие цепких когтей, а также хорошо развитые зрительная система и вестибулярный аппарат. Многие виды лазают по стволам либо лазают или бегают по веткам Некоторые обезьяны способны к брахиации – передвижению при помощи передних конечностей, поочередно закидываемых вперед, тогда как тело висит в воздухе В воздухе. Хотя представители ряда различных групп, например летучие рыбы и летяги, способны к планированию, главной формой передвижения в воздухе служит истинный полет, наблюдаемый у насекомых, птиц и летучих мышей. У птиц наиболее распространенный тип полета – это машущий полет, при котором птица ритмично поднимает и опускает крылья. При этом внутренние части крыльев обеспечивают главным образом создание подъемной силы, преодолевающей силу тяжести, а наружные части создают пропеллирующий эффект, толкая птицу вперед (Hein-roth, Heinroth, 1958). Под землей. Роющими или полуроющими называют виды, которые всю свою жизнь или большую ее часть проводят под землей. У таких животных обычно имеются многочисленные адаптации (малая величина глаз и ушных раковин, менее густая шерсть и др), уменьшающие трение при передвижении под землей. На поверхности земли. Существует много разных типов передвижения животных по земле У ряда различных млекопитающих наблюдается хождение на двух ногах, наиболее характерное для некоторых кенгуру и грызунов, а также для человека Такие животные, как кенгуру и кенгуровые крысы, передвигаются прыжками Формы локомоции у большинства видов, передвигающихся на четырех конечностях, довольно стереотипны. При ходьбе конечности переставляются в таком порядке, чтобы всегда сохранялась опора в виде треугольника, создаваемого тремя конечностями. При большей скорости движения, например при рыси и галопе, устойчивость снижается (Gray, 1953). Гну и львы способны бежать со скоростью до 80 км/ч, а гепарды – даже до 110 – 120 км/ч (Bourliere, 1964). ПИТАНИЕ И ДЫХАНИЕ Потребление пищи, питье воды и дыхание имеют решающее значение для выживания самых разнообразных животных. Потребление пищи Стратегии питания. Животных можно разделить по их главной стратегии питания на пять основных групп, фильтрующие воду, паразиты, растительноядные, плотоядные и всеядные (Maier, Maier, 1970). Все фильтрующие организмы живут в воде и добывают себе пищу, отцеживая ее из воды Движение воды относительно тела животного достигается либо путем того или иного нагнетающего действия, либо за счет плавания Паразиты, как правило, живут целиком за счет организма-хозяина; некоторые из них (например, лентецы) живут в течение долгого времени, не убивая своего хозяина, а другие довольно быстро вызывают его гибель. Растительноядные питаются растениями, их корой, листьями, семенами, плодами и корнями. Живущий в Австралии коала высо-коспециализирован в отношении пищи и питается исключительно листьями эвкалипта У жвачных имеется четырехкамерный желудок, обеспечивающий более полное переваривание пищи. Плотоядные питаются главным образом другими животными Как среди беспозвоночных, так и среди позвоночных можно найти виды с самыми разнообразными типами питания. Многие животные всеядны, они поедают всевозможные виды пищи. У таких животных стратегии питания наименее специализированы Например, бродячие муравьи уничтожают почти все, что попадается на их пути в тропическом лесу Весьма разнообразные формы всеядности можно встретить у грызунов (Landry, 1970) Даже шимпанзе, по-видимому, всеядны, причем для них характерен удивительно высокий уровень организации охоты (Teleki, 1973а, Ь). Формы поведения, связанные с питанием. Из таких форм поведения мы рассмотрим хищничество, запасание пищи, разного рода манипуляции с нею и регулирование ее потребления. Хищничество. Некоторые виды, например пауки, строящие паутину, богомолы и муравьиные львы, пассивно поджидают добычу, тогда как большинство видов активно охотится. Пойманную добычу хищник хватает, душит, сжимает, отравляет, обездвиживает, оглушает или поедает живьем. Диапазон приспособлений к хищничеству поистине велик. Последовательность действий хищного млекопитающего можно разбить на приближение к жертве, ее умерщвление и поедание (Ewer, 1968). Домашние кошки подкрадываются к своей жертве, прижимаясь к земле, следя за каждым ее движением и используя короткие перебежки, чтобы все ближе и ближе подойти к ней, а затем совершить нападение. В отличие от этого гепарды приближаются к жертве, преследуя ее в открытую, и используют свою исключительную скорость бега лишь в том случае, когда жертва обращается в бегство. Млекопитающие часто убивают свою жертву, прокусывая ей затылок. Совершаемые при этом действия различаются в зависимости от вида хищника и от обстоятельств. Иногда хищник сначала схватывает жертву в лапы или прижимает ее к земле. Для многих видов, в частности для собачьих, характерно ритуализо-ванное «умерщвление встряской»: схватив жертву в зубы, хищник размахивает ею из стороны в сторону (Eisenberg, Leyhausen, 1972). Большинство хищников сначала поедает голову жертвы, а затем постепенно съедает ее всю в направлении от головы к заднему концу тела. Как показал Лейхаузен, ориентация жертвы обусловливается прежде всего осязательными стимулами – расположением шерсти или оперения. Поедание крупной добычи часто начинается с брюха или паховой области (Ewer, 1968). Насытившись, хищник обычно прекращает поиски пищи и охоту. Лишь в редких случаях он убивает больше того, что способен съесть;, это может произойти в условиях, когда плотность популяции жертвы очень велика и поэтому усилия, затрачиваемые на ее поиски, минимальны (Kruuk, 1972). Запасание пищи. Поскольку количество пищи в разное время года неодинаково, многие животные, особенно грызуны, делают запасы. Существуют две главные формы запасов: сконцентрированные в специальных «кладовых» и рассеянные по разным местам отдельными кучками (Ewer, 1968). Кладовые обычно устраиваются в самой норе, часто в специальной камере, а при рассеянном хранении запасов они могут быть разбросаны по значительно более обширной площади и находиться довольно далеко от дома. Встречаются и промежуточные стратегии. В норах прерийных полевок – небольших грызунов, обитающих в полях, – находили запасы хорошо сохранившихся семян объемом до 9 – 10 литров (Jameson, 1947). У многих видов, запасающих корм, имеются обширные защечные мешки, помогающие животным переносить корм к месту его хранения. Запасание корма у грызунов можно вызвать в лабораторных условиях; изучалось влияние, которое оказывают на эту форму поведения лишение пищи, индивидуальный опыт в ранний период жизни, различные физиологические воздействия и другие подобные переменные. Манипуляции с пищей. Характер манипуляций, производимых с пищей до ее потребления, сильно варьирует у разных видов и зависит от типа пищи. Так, например, калан ложится в воде на спину, кладет себе на грудь плоский камень и вскрывает раковины моллюсков, ударяя ими об этот камень. Другие животные используют для разного рода манипуляций передние лапы и пасть. Для развития многих типов манипуляций с пищей, по-видимому, достаточно лишь минимального опыта..Раскалывая орехи, белки прогрызают в скорлупе одну или две бороздки, расположенные определенным образом. Эйбл-Эйбесфельдт (Eibl-Eibes-feldt, 1963) вырастил в неволе белок, которым не приходилось прежде раскалывать орехи, и когда животные впервые приступили к этому, разгрызающие движения в общем не отличались от обычных, хотя были довольно плохо ориентированы. Регуляция потребления пищи. Животные не кормятся непрерывно. По мере того как проходит все больше времени после кормежки, особи многих видов становятся более активными и начинают особенно живо реагировать на стимулы, ассоциированные с пищей. Потребление пищи вовремя прекращается; у большинства животных проблема ожирения возникает редко. У млекопитающих потребление пищи регулируется, по-видимому, в результате взаимодействия двух ядер гипоталамуса – латерального и вентромедиального (Keesey, Powley, 1975). В отличие от этого у падальных мух потребление пищи регулируется в основном периферически (т. е. меньше зависит от центральной нервной системы): его инициируют периферические рецепторы, а тормозят импульсы, поступающие от растянутой передней кишки (Dethier, 1967). Потребление воды Вода совершенно необходима для самых разнообразных жизненных процессов, в том числе для пищеварения, выделения и терморегуляции. Многие животные должны активно поглощать воду как таковую, тогда как другим достаточно воды, извлекаемой в процессе метаболизма из пищевых веществ. Разные животные используют при питье различные движения. Собаки и волки принимают стереотипную позу и лакают воду языком, тогда как корова всасывает воду ртом, а язык играет у нее лишь пассивную роль. У большинства грызунов наблюдается довольно стереотипное слизывание воды. Слоны, конечно, используют хобот, чтобы насосать воду и перенести ее в рот (Bunnell, 1973). Дыхание Потребление кислорода путем дыхания – явление столь универсальное, что его нередко упускают из виду. Практически все животные обладают тем или иным механизмом, при помощи которого свежий воздух поступает в организм, а использованный выводится наружу. Дыхание становится проблемой первостепенной важности у водных млекопитающих. Дыхательные системы у них обладают удивительными приспособлениями, позволяющими этим животным нырять на большие глубины и оставаться под водой дольше, чем это могут другие млекопитающие. По имеющимся данным, ондатра и морской слон способны оставаться под водой в течение 12 минут, тогда как кит-бутылконос может погружаться на 120 минут (Bourliere, 1964). ТЕРМОРЕГУЛЯЦИЯ Животные способны существовать только в определенном диапазоне температур. Регуляция температуры тела – важнейший аспект приспособления к среде, особенно в суровых условиях пустыни или Арктики. Птицы и млекопитающие – гомойотермные животные; у них имеются физиологические механизмы, поддерживающие температуру тела на относительно постоянном уровне, несмотря на резкие колебания температуры среды. В отличие от этого беспозвоночные, рыбы, амфибии и рептилии пойкилотерм-ны. – температура тела у них колеблется в зависимости от внешней температуры. Многие формы поведения по крайней мере частично связаны с терморегуляцией. Для многих видов характерны сезонные миграции, назначение которых в том, чтобы сводить к минимуму воздействие неблагоприятных температур в определенное время года. Более кратковременная регуляция температуры тела достигается путем значительных ежедневных перемещений из затененных участков в освещенные или из норы наружу и обратно. В лабораторных условиях животное может научиться выполнять произвольные действия, если изменения температуры используются в качестве подкрепления (см., например, Wright, Meyer, 1969). Многие виды переживают суровые зимы, впадая в зимнюю спячку. При этом животное прекращает всякую активность, а температура тела, частота сердечных сокращений и уровень метаболизма снижаются у него до минимума. Некоторые виды лягушек, крокодилов и рыб реагируют аналогичным прекращением активности и на крайне высокие температуры – это так называемая летняя спячка. ПОИСКИ УБЕЖИЩ Большинство видов предпринимает поиски каких-либо убежищ, чтобы укрыться от резких колебаний температуры, осадков и хищников. Иногда животное просто забирается в пещеру, расселину или на дерево, что не приводит к сколько-нибудь существенному изменению среды. Однако во многих других случаях оно строит весьма сложные гнезда или норы, что влечет за собой значительные изменения внешних условий. Постройки бывают относительно постоянными, как, например, плотины бобров, или временными, как спальные гнезда шимпанзе, в которых они обычно проводят лишь одну ночь. У многих видов устройство гнезда тесно связано с размножением: когда близится время появления потомства, они приступают к постройке гнезда или значительно расширяют уже имеющееся гнездо. Среди беспозвоночных постройка убежищ наиболее ярко выражена у насекомых. У таких видов, как одиночные осы, каждая отдельная самка роет норку и запасает в ней корм. Однако у многих других видов гнезда представляют собой весьма сложные постройки, и в них обитают целые сообщества. Примером служат высокие сооружения термитов и гнезда пчел. Структура сообщества у разных видов пчел различна, но в них непременно имеется одна матка и много рабочих особей. У медоносной пчелы отличительная черта деятельности рабочих особей по постройке и содержанию гнезда – функциональная специализация. Сооружением сотов, кормлением личинок, уборкой ячеек, приготовлением меда, охраной входа и сбором пыльцы и нектара занимаются разные особи. При этом функции каждой отдельной рабочей пчелы изменяются в течение ее жизни: она начинает с чистки ячеек и заканчивает сбором пыльцы и нектара (Lindauer, 1961). Выбор места для нового гнезда у медоносной пчелы – процесс весьма интересный. В конце весны матка и около половины рабочих особей оставляют старое место дочерям матки и образуют рой на небольшом расстоянии от него. Пчелы остаются в этом рое до тех пор, пока не будет выбрано новое место. Пчелы-разведчицы вылетают из роя для обследования многочисленных мест, потенциально пригодных для устройства гнезда. Возвращаясь в рой, они исполняют «танец», содержащий указания о том, где находятся эти места (см. гл. 5). Интенсивность танца варьирует в зависимости от качества данного места. Особое значение придается, по-видимому, его величине и защитным возможностям. Танцующие пчелы вербуют новых разведчиц. На основании интенсивности танцев и реакции новых разведчиц рой «принимает решение»: в конце концов преобладающая часть разведчиц указывает при помощи танца одно определенное место, и тогда рой снимается и направляется туда (Lindauer, 1961). У грызунов поведение, связанное с поиском или постройкой убежища, принимает весьма разнообразные формы. Бобры строят однокамерные норы, или хатки, в которых обитают пара взрослых особей и два их последних выводка. Древесные крысы собирают множество прутиков или веточек, из которых строят обширные шалаши. Подробно изучена постройка гнезд лабораторными крысами и домовыми мышами. Оба вида устраивают гнезда в форме кубков или чаш, используя вату, бумагу, тряпки и другие подобные материалы; иногда гнезда снабжаются крышей. Шимпанзе, орангутаны и гориллы строят себе «спальные» гнезда на деревьях. ИЗБЕГАНИЕ ХИЩНИКОВ Поскольку большинство видов служит добычей по меньшей мере для нескольких других видов, избегание хищников имеет важное значение для выживания и размножения. Главные способы защиты от хищников – это укрывание от них, предупреждение особей своего вида, наличие предостерегающих признаков, бегство и активное сопротивление. Укрывание Многие животные скрываются от хищников в убежищах – норах, щелях и шалашах. Кроме того, укрытию от хищника может способствовать внешний вид самого животного. Покровительственная окраска, благодаря которой животное сливается с фоном, встречается у представителей почти всех таксономических групп. Много особенно ярких примеров можно найти у насекомых, среди которых есть формы, сходные с листьями, веточками или даже с птичьим пометом. Часто покровительственная окраска сочетается с особым поведением: животное располагается по отношению к окружающему определенным образом, обычно сохраняя неподвижность.
Бегство Быстрота и проворность служат наилучшим и, вероятно, наиболее распространенным средством спасения от хищников. Многие виды, спасаясь бегством, дополняют свои локомоторные движения демонстрационным поведением с целью отвлечь внимание потенциального хищника или испугать его. Другие, напротив, затаиваются, чтобы уменьшить вероятность нападения (см. Galiup, 1974). Активное сопротивление В качестве последнего средства защиты жертва может оказать хищнику активное сопротивление При этом она может ударить хищника, схватить его или укусить. Скунсы и многие виды членистоногих, например многоножки, выделяют химические вещества, отпугивающие хищников. Другие животные защищаются от хищников своими плотными или ядовитыми покровами, твердой раковиной или такими выростами, как шипы и иглы. СОН Определение сна может быть основано на поведенческих или электрофизиологических критериях или же на сочетании тех и других. В аспекте поведения сон – это период длительного отсутствия активности, для которого обычно характерны четыре признака: 1) часто – суточный (или приливной) ритм; 2) обычно – повышение порога различных реакций; 3) часто – приуроченность к определенным местам (обычно какое-либо надежное убежище); 4) характерная для данного вида поза. Под такое определение подпадает сон, наблюдаемый у широкого круга животных, в том числе у насекомых, моллюсков, рыб, амфибий, рептилий, птиц и млекопитающих (Meddis, 1975). Электрофизиологические сведения относительно -сна дает электроэнцефалограмма (ЭЭГ). Это запись активности головного мозга («мозговых волн»), получаемая с помощью электродов, которые либо прикладывают к коже головы снаружи, либо вживляют в мозг. Электрофизиологические данные о сне получены только для птиц и млекопитающих. У этих животных можно различить две главные фазы сна: медленноволновой сон и парадоксальный сон (рис. 4.2). Для медленноволнового сна характерны высокоампли-тудные медленные колебания, появляющиеся в ЭЭГ. Об общем расслаблении свидетельствует падение мышечного тонуса, отмечаемое, например, при регистрации электрической активности шейных мышц (рис. 4.2).
Во время сна периодически отмечаются резкие изменения в ЭЭГ и характере мышечной активности. Хотя, судя по внешним признакам, животное продолжает спать, его ЭЭГ становится «десинхронизированной», с быстрыми низкоамплитудными колебаниями. Тонус скелетных мышц падает ниже уровня, обычного для медленноволнового сна. Поскольку животное кажется спящим, в то время как его ЭЭГ сходна с ЭЭГ при бодрствовании, эту фазу часто называют парадоксальным сном. Одна из самых характерных особенностей парадоксального сна – появление быстрых движений глаз (поэтому сон такого типа называют также БДГ-сном). Парадоксальный сон, во всяком случае у человека, связан со сновидениями. Периоды парадоксального сна возникают примерно каждые 9 минут у домовой мыши, каждые 50 минут у обезьян и детей и примерно каждые 90 минут у взрослых людей. ПОДДЕРЖАНИЕ ЧИСТОТЫ ТЕЛА Многим животным для здоровья и благополучия необходимо поддерживать поверхность тела в чистоте и не допускать заражения эктопаразитами. У большинства млекопитающих чистка шерсти (груминг) составляет одно из каждодневных занятий. Часто она производится сразу после еды. Многие грызуны «купаются в песке» – изгибают тело и трутся боками и брюшком о песок, чтобы удалить с шерсти посторонние частицы и избыток жирового материала, выделяемого кожными железами (см. Borchelt, Gris-wold, Branchek, 1976). У многих птиц в различных ситуациях можно наблюдать чистку оперения. Иногда поддержание чистоты тела обеспечивают представители другого вида. Многие виды «рыб-чистильщиков» приспособлены к тому, чтобы удалять паразитов с покровов гораздо более крупных рыб. Некоторые из них выбирают паразитов даже из ротовой полости крупных хищных рыб, которые их при этом не съедают. У приматов груминг не только служит для содержания шкуры в чистоте, но и представляет собой важный фактор поддержания структуры сообщества. Груминг превратился у них в тонкое искусство и у некоторых видов занимает ежедневно довольно много времени. У лори и других лемуров из нижних клыков и резцов образовались специальные «зубные гребешки», используемые для чистки шерсти, роль груминга во взаимоотношениях между особями, быть может, наиболее ярко выражена в случае матери и детеныша и самца и самки (где груминг может служить прелюдией к копуляции); он играет также большую роль в установлении иерархических взаимоотношений в сообществе (Jolly, 1972). Шимпанзе в неволе иногда используют деревянные палочки для ковыряния в зубах друг у друга (McGrew, Tutin, 1973). Шимпанзе-«дантист» пользуется палочками для удаления остатков пищи с зубов «пациента». ВЫДЕЛЕНИЕ Особенности мочеиспускания и дефекации у разных видов представляют гораздо больший интерес для исследователя поведения животных, чем это может показаться на первый взгляд. Прежде всего для многих видов характерны определенные позы, принимаемые при мочеиспускании и дефекации. У собак, например, наблюдается половой диморфизм в позе при мочеиспускании: взрослые суки при этом приседают, а кобели поднимают заднюю ногу Степень внимания, уделяемого выбору места для мочеиспускания или дефекации, по-видимому, варьирует в зависимости от характера гнездового участка и его постоянства. Приматов, у которых стадо бродит по территории обитания, ночуя всякий раз в разных местах, мало заботит дальнейшая судьба мочи или кала. У видов с более постоянным гнездовым участком, например у многих хищных, этот участок должен содержаться.в чистоте, а поэтому контроль за мочой и экскрементами строже. Домашние кошки обычно зарывают экскременты в землю. У многих млекопитающих с защищенными гнездовыми участками матери поедают мочу и экскременты своих детенышей, часто до того времени, пока детеныши не начинают самостоятельно передвигаться. Это не только поддерживает чистоту гнезда, но и уменьшает вероятность того, что детеныши будут обнаружены хищником по запаху мочи и кала. Чайки, гнездящиеся на земле, удаляются для дефекации на некоторое расстояние от гнезда, а у моевок, обитающих на скалах, дефекация происходит просто через край гнезда. Характерный для данного вида процесс мочеиспускания и дефекации может развиваться медленно. У многих млекопитающих, например у морских свинок, матерям приходится стимулировать мочеиспускание и дефекацию у новорожденных детенышей, вылизывая им аногенитальную область. Развитие разных поз при мочеиспускании у собак, по-видимому, регулируется гормонами (Beach, 1974). Моча и экскременты могут играть важную роль в общении между животными, особенно в тех случаях, когда оно основано на обонятельных сигналах. У многих видов моча и экскременты используются в качестве меток, указывающих на то, что данный участок занят. Поднимание задней ноги у домашних собак и обливание предметов направленной назад струёй мочи у котов, по-видимому, обеспечивает разбрызгивание мочи на достаточную высоту. Бегемоты при дефекации производят быстрые движения хвостом, что приводит к разбрасыванию навоза по большой площади. Химические сигналы, исходящие от продуктов выделения, играют также важную роль в регуляции репродуктивной активности. Они могут блокировать развитие беременности, вызывать течку у самок в диэструсе или ускорять половое созревание (Вгоп-son, 1971) (см. гл. 10). У таких животных, как шиншиллы, морские свинки, дикобразы и кролики, в процессе ухаживания часто наблюдается энурина-ция – испускание самцом мочи, направляемой в сторону самки. Этот акт не входит в обычную последовательность действий при ухаживании и, возможно, отражает либо «фрустрацию», либо «неприятие» (Ewer, 1968). Многие сравнительные психологи пытаются оценивать уровень «эмоциональности» животных. Одним из используемых при этом показателей служит количество экскрементов, выделяемых животным, которое поместили в новую для него ситуацию. ИССЛЕДОВАТЕЛЬСКОЕ ПОВЕДЕНИЕ Для большинства видов животных знакомство с окружающей средой часто приносит большую пользу, облегчая выживание и размножение. Систематически исследуя свой участок, животное получает представление о местонахождении пищевых и других ресурсов, потенциальных брачных ^партнеров и мест, где можно укрыться от хищников. Поэтому нередко можно видеть, как животные, которые досыта наелись и напились и не находятся в состоянии готовности к спариванию, тем не менее обследуют свою территорию. Хотя лишение воды или пищи, эструс и другие факторы могут способствовать исследовательской активности, они не необходимы для ее проявления. Формам исследовательского поведения посвящено много работ зоопсихологов. Обследование предметов Чтобы изучать обследование животными новых предметов, можно просто вносить такие предметы в клетку. Гликмен и Срод-жес (Glickman, Sroges, 1966) изучали реакцию более чем 300 животных, принадлежащих к более чем 100 видам, на куски дерева, стальные цепи, деревянные палочки, резиновые трубки и шарики из мятой бумаги. Результаты их экспериментов представлены на рис. 4.3. Приматы и хищные проявляли к обследованию предметов гораздо больший интерес, чем животные с менее развитым головным мозгом; наименее активными в этом отношении оказались рептилии. Характерно то, что такая деятельность была выражена в наивысшей степени в начале 6-минутных тестов, а к концу теста постепенно снижалась.
Другой метод состоит в том, что новый предмет закрепляют в небольшой нише и дают животному возможность приближаться к нему. Для регистрации числа таких приближений можно использовать фотоэлемент («электрический глаз»). Новое как подкрепление Как было установлено, животные научаются выполнять разнообразные задачи в условиях, когда единственным подкреплением служит возможность проводить исследовательскую деятельность. Например, крысы учатся находить выходы из лабиринтов, когда подкрепление состоит в возможности обследовать новую сложную среду. Макаки-резусы нажимают на рычаг, открывающий окно, получая в качестве подкрепления возможность увидеть через это окно интересные вещи. Наибольший интерес они проявляли к другим обезьянам, затем шли (в порядке ослабления интереса) электрические поезда, пища и пустая камера (Butler, 1960). Сложность стимула, по-видимому, играет решающую роль в вызывании исследовательского поведения у млекопитающих (Walker, 1964). Неофобия В некоторых условиях животные избегают новых предметов – явление, получившее название неофобии (Barnett, 1963). Подобная реакция особенно обычна при внезапном изменении привычной обстановки. Если диким серым крысам, выращенным в лаборатории и привыкшим брать корм из проволочной корзинки, прикрепленной к задней стенке их клетки, предложить корм в корзинке у передней стенки, то они могут отказываться от пищи в течение нескольких дней. Такое поведение – пример неофобии. Ручные лабораторные крысы начинают обследовать корзинку на новом месте раньше, и ее перенос меньше влияет на их ежедневный прием пищи. Создается впечатление, что неофобическая реакция у этих крыс ослаблена в результате приручения (Barnett, 1963; Mitchell, 1976; Barnett Cowan, 1976). ИГРА У разных видов животных, особенно у млекопитающих, наблюдаются типы поведения, которые можно смело назвать игрой. Детеныши хищников, например домашней кошки, подолгу возятся с такими предметами, как мяч, – приводят его в движение, прыгают на него, катают по полу и схватывают. Волчата и лисята бешено крутятся, пытаясь схватить себя за хвост. Случалось наблюдать за игрой выдр, которые скатывались по снежному склону (Bourliere, 1964). В плане общественного поведения часто можно видеть игры детенышей хищных и приматов, гоняющихся друг за другом, затевающих шуточные драки и беготню. Большинство наблюдателей согласились бы, что все это – формы игрового поведения, однако дать определение игры оказалось трудной задачей. Все виды игрового поведения, по-видимому, возникают в такое время, когда нет необходимости ни в каких других формах поведения, существенных для выживания, таких, как питание или спасение от хищников. Игровое поведение как бы лишено «серьезности», присущей большинству других форм поведения. Игра, очевидно, «доставляет удовольствие» ее участникам. В плане общественного поведения одно животное может вызывать другое на игру и подавать сигнал о том, что «все последующее – это игра» (Bekoff, 1974a). Игра чаще наблюдается у детенышей, чем у взрослых особей. Различные исследователи, изучавшие игру, рассматривали ее либо с функциональной, либо со структурной точки зрения (Fagen, 1974). Лоизос (Loizos, 1966) сделала попытку охарактеризовать игру в структурном плане. Она отметила, что игра в большинстве случаев сопряжена с перестройкой стереотипных последовательностей движений, наблюдаемых у взрослых особей, например при ловле добычи или половой активности. Такого рода перестройки можно подразделить на шесть типов: 1) может быть изменена последовательность движений; 2) отдельные двигательные акты, входящие в последовательность, могут быть преувеличены; 3) некоторые движения, входящие в последовательность, могут многократно повторяться; 4) нормальная последовательность может остаться незавершенной, т. е. закончиться раньше, чем обычно, в результате перехода к посторонним действиям; 5) некоторые движения могут быть и преувеличены., и многократно повторены; 6) отдельные движения, входящие в последовательность, могут оставаться незавершенными. Кроме того, могут перемешиваться акты, обычно связанные с совершенно разной мотивацией. Высказывалось много предположений относительно роли игры в жизни животных. Поскольку игра особенно характерна для детенышей млекопитающих, большинство исследователей считает, что она имеет какое-то значение для развития нормального поведения взрослых особей. Игра, возможно, позволяет детенышам практиковаться в выполнении двигательных актов и общественных взаимодействиях, которые будут необходимы во взрослом состоянии, и совершенствовать их. Она, несомненно, служит тренировкой и обогащает информацию об окружающей среде. Исследования относительно функций игры очень трудны, но крайне необходимы ИСПОЛЬЗОВАНИЕ ОРУДИЙ Самые различные животные используют орудия в разнообразных ситуациях, в частности для добывания пищи и для защиты Элкок (Alcock, 1972) дает превосходный обзор использования орудий, связанного с питанием, и предлагает следующее определение (р. 464): «Использование орудий состоит в манипуляциях с ка ким-либо неодушвленным предметом, созданным вне собственно-ю организма животного и повышающим эффективность его действий, направленных на изменение положения или формы какого-либо другого предмета». Если принять это определение, то такие формы поведения, как взаимодействия между особями и ловля добычи с помощью паутины, нельзя считать использованием орудий. Примеров использования животными орудий можно привести великое множество. Мы уже говорили о применении морскими выдрами камней для вскрытия раковин моллюсков и об использовании шимпанзе деревянных палочек для чистки зубов друг другу. Рыбы-брызгуны направляют струйки воды на находящуюся на суше жертву, с тем чтобы сбросить ее в воду, где ее можно будет изловить. Разные виды птиц используют прутики или колючки кактусов для извлечения насекомых из коры или других недоступных мест. Сообщалось о том, что некоторые птицы разбивают яйца других птиц, сбрасывая на них камни, а затем выедают содержимое. Ван Лавик-Гудолл (Van Lavick-Goodall, 1967) описывает у шимпанзе два интересных примера изготовления орудий (обработка предметов, которые затем используются в качестве орудий). В одном случае ветви или стебли растений сначала очищаются от боковых веточек и вводятся в гнезда термитов; прилипших к такому прутику термитов обезьяна поедает. В другом случае шимпанзе пережевывают листья, сжимают их в комки и используют в качестве губки, добывая из древесных дупел питьевую воду. Ясно, что формы поведения, связанные с использованием орудий, возникали независимо много раз и что они часто представляют собой лишь незначительные модификации в существующем репертуаре поведенческих актов соответствующих животных. БИОЛОГИЧЕСКИЕ РИТМЫ Одна из самых поразительных особенностей поведения – это его организация во времени. Строгая цикличность, наблюдаемая в поведении буквально всех видов животных, позволяет предсказывать, когда проявится та или иная его форма. Временные масштабы биологических ритмов могут быть совершенно различными. Периодические цикады выводятся в данной области, буквально наводняя ее, один раз в каждые 13 или 17 лет. Циклы колебаний реактивности нервных клеток занимают несколько миллисекунд. Существуют годичные циклы, приливные циклы, циклы питания, астральные и менструальные циклы и т. д. В сущности, можно рассматривать почти всю науку о поведении животных как изучение ритмов, лежащих в основе различных форм поведения в разное время, и факторов, регулирующих эти ритмы. Много внимания уделялось изучению ритмов, периодичность которых составляет около суток. Такие ритмы называются цир-кадными (от лат. circa – около и dies – день). Поведение многих птиц и млекопитающих, по-видимому, связано с циркадными ритмами. Дневные виды обычно спят ночью, а активны при дневном свете, тогда как ночные виды активны ночью, а спят днем Сумеречные виды активны на рассвете, в сумерки или и в тот и другой период. Для изучения циркадных ритмов часто используют бег в колесе, который легко оценить количественно. Животное помещают внутрь небольшого колеса, надетого на горизонтальную ось. К колесу примыкает небольшая клетка, так что животное может входить в колесо и выходить из него. Ряд рычагов и счетчиков позволяет регистрировать число поворотов колеса, производимых животными в единицу времени. Если грызуна, ведущего ночной образ жизни, например хомяка, поместить в такое колесо и чередовать смену света и темнота каждые 12 часов, то у животного наблюдается упорядоченная картина активности: оно начинает бегать в колесе примерно в то время, когда свет выключается, и заканчивает ко времени включения света. Такой ритм называется экзо-генным – он сопряжен с каким-либо периодическим процессом, происходящим во внешней среде, в данном случае с циклическим чередованием света и темноты. Периодический внешний стимул, с которым синхронизирован биологический ритм, называют «датчиком времени» или «синхронизатором» (см. Rusak, Zucker, 1975).
Одна из главных проблем в изучении циркадных ритмов состоит в том, чтобы выяснить, являются ли все они экзогенными (зависящими от периодического воздействия какого-то датчика времени) или эндогенными (для поддержания которых не требуется никаких регуляторных внешних стимулов). Для установления: эндогенпости ритма животных обычно содержат в условиях непрерывного освещения или постоянной темноты, регистрируя при этом их активность или другие ритмы В таких условиях у многих животных наблюдается продолжение обычных для них периодических колебаний активности, хотя циклы могут быть несколько длиннее или короче суток (рис 45 и 46). Согласно «правилу Ашофа», направление и степень такого отклонения от строгого 24-часового цикла зависит от интенсивности освещения и от того, в какое время активно данное животное в природных условиях – днем или ночью (Marler, Hamilton, 1966). Хотя известны исключения из этого правила, в общем оно оказалось полезным. Продолжение данного ритма в условиях постоянного освещения или темноты позволяет думать, что он является эндогенным, хотя нельзя исключить, что существует какой-то невыявленный фактор, действующий как датчик времени. Ряд различных данных говорит о том, что ритмы могут быть подлинно эндогенными (Marler, Hamilton, 1966). /. Эксперименты с переносом. Если по видимости эндогенный ритм на самом деле подчиняется какому-то неустановленному стимулу, связанному с вращением Земли, то после перевозки животного на другую долготу этот ритм должен тотчас же измениться Обычно этого не происходит; у животных, перевезенных таким образом без изменения прочих условий, сохраняются первоначальные ритмы (Трудности приспособления к новому ритму хорошо знакомы всем, кто совершал дальние перелеты на реактив ных самолетах) 2. «Остановка часов». Охлаждая животных почти до замерзания, можно, по существу, остановить ход предполагаемых «биологических часов» организма. После согревания животного цикл •его активности возобновляется, но уже не совпадает по фазе ни с каким внешним циклическим процессом, который мог бы служить датчиком времени 3. Отклонение от 24-часового ритма Сам факт, что периодичность большинства «независимых» циклов несколько короче или длиннее 24 часов, заставляет сомневаться в их экзогенной регуляции Трудно представить себе стимул, который изменялся бы столь упорядоченным образом и при этом был бы чувствителен к интенсивности света в камере, так что мог бы подчиняться пра-вилу^Ашофа. Концепцию об экзогенной регуляции циркадных ритмов отстаивал Ф. Браун (см, например. Brown, 1972). Онтогенез многих циркадных ритмов, по-видимому, сильно зависит от генетических факторов Так, например, у всех ящериц, которых выращивали при разных необычных режимах света и темноты, а затем помещали в постоянные условия, наблюдались сходные циркадные ритмы (см Marler, Hamilton, 1966). КРАТКИЕ ВЫВОДЫ Целью этой главы было вводное описание разнообразных форм поведения, позволяющих отдельным организмам приспособляться к своей среде и выживать в ней Были рассмотрены такие функции, как локомоция, питание, терморегуляция, поиски убежищ, избегание хищников, сон, поддержание чистоты тела, выделение,, исследовательская активность, игра, использование орудий и биологические ритмы. Даже такой беглый обзор ясно показал разнообразие форм индивидуального поведения и тех способов, которыми они обеспечивают выживание. Ни одна форма поведения не существует в. отрыве от других; все они взаимодействуют друг с другом, и в результате мы имеем организм, способный к выживанию и размножению в определенной среде. Глава 7 ГЕНЕТИКА ПОВЕДЕНИЯ Ученые давно интересовались и теориями, и фактическими данными относительно роли наследственных факторов в поведении. Подобный интерес можно проследить со времен древних греков, затем у Леонардо, а далее у Монтеня, Дарвина, Гальтона, Иеркса и Трайона (McClearn, DeFries, 1973). Однако как отдельная дисциплина генетика поведения совсем еще молода. Заслуга выделения ее как четко определенной области исследования принадлежит Джону Фуллеру и У. Р. Томсону, которые в 1960 г. опубликовали свою монографию «Генетика поведения» («Behavior genetics»). В последние 20 лет изучение влияния генетических факторов на поведение быстро продвигалось вперед. Поскольку разные исследователи интересуются разными проблемами, работы в этой области удобно разбить на шесть групп, соответствующих следующим вопросам (по Thiessen, 1972, видоизменено): 1. Влияют ли различия в генотипе на исследуемое поведение? 2. Если влияют, то сколько генов к этому причастно? 3. Какая доля наблюдаемой изменчивости поведения определяется генетическими и какая – средовыми факторами? 4. Какие онтогенетические и физиологические процессы опосредуют влияние генов на поведение? 5. Что может дать генетический анализ для выяснения возможной адаптивной роли данной формы поведения? 6. Как взаимодействуют гены и внешняя среда в детерминации данного поведения? В настоящей главе будут рассмотрены эти шесть фундаментальных вопросов, методы, которые используются при их изучении, и некоторые уже найденные ответы. При обсуждении данных по генетике поведения нам придется использовать ряд основных генетических понятий. Часть из них уже была разъяснена в главе 3; возможно, читателю полезно будет еще раз вернуться к этому месту, прежде чем идти дальше.
Межлинейные различия Основной принцип использования межлинейных различий для решения интересующего нас вопроса очень прост. На развитие поведения влияют два главных фактора – гены и среда. Стратегия исследования состоит в том, чтобы найти организмы, различающиеся по генотипу, и, поддерживая постоянство внешних условий, определить, влияют ли на поведение генетические факторы. Главная трудность генетического исследования связана с тем, что гены нельзя наблюдать непосредственно. Для того чтобы делать выводы относительно генотипа, приходится применять косвенные методы. Как же в таком случае мы можем найти организмы с различным генотипом? Многие исследователи используют для этого инбредные линии. В инбредной линии все особи фактически гомогенны, т. е. генетически идентичны, и гомозиготны по всем локусам. Инбредные линии домовой мыши (Mus musculus) – наиболее -популярного объекта в исследованиях по генетике поведения млекопитающих – выводят путем скрещивания братьев с сестрами на протяжении по меньшей мере 20 поколений. Из генетически гетерогенной популяции берут брата и сестру и получают от них потомство. Брата и сестру из этого потомства снова скрещивают между собой для получения следующего поколения, и т. д. С каждым поколением данная линия безвозвратно теряет некоторые аллели, и примерно к 20-му поколению она приближается к полностью изогенному состоянию. Отметим, что в этом случае животных для скрещивания подбирают на основе их генетического родства – в отличие от селекции на определенные признаки (анатомические, физиологические или поведенческие), о которой речь пойдет позже. В значительной мере благодаря большой программе, осуществленной в Джексоновской лаборатории (штат Мэн, США), в настоящее время исследователи располагают большим числом инбредных линий животных. Каждая из них получила свое обозначение в соответствии с международной системой (например, C57BL/6J). Если животные разных инбредных линий, выращенные в идентичных условиях среды, обнаруживают существенные различия, то весьма вероятно, что эти различия имеют генетическую основу. Подобные результаты были получены для целого ряда особенностей поведения, включая активность в «открытом поле» (Thompson, 1953), агрессивное поведение (Southwick, Clark, 1968), половое поведение (McGill, 1962), постройку гнезда (Lynch, Heg-mann, 1972) и научение (Wahlsten, 1972). Фактически даже труднее найти линии, которые не отличались бы друг от друга, чем линии, между которыми есть различия. Спрот и Стаатс (Sprott, Staats, 1975) составили полную библиографию (1222 названия) работ по поведению с использованием генетически охарактеризованных мышей. Перекрестное воспитание. Хотя межлинейные различия указывают на генетический эффект, не исключено, что эти различия обусловлены какими-то тонкими средовыми факторами. Один из способов контроля таких факторов – эксперимент с перекрестным воспитанием. Пометы новорожденных мышат, принадлежащие к линиям с разным поведением во взрослом состоянии, меняются местами, так что матери одной линии воспитывают детенышей другой, и наоборот. Контрольные животные воспитываются либо собственными матерями, либо другими самками той же линии. Один эксперимент такого рода был проведен Саутвиком (Southwick, 1968). Линии A/J и CFW сильно различались по агрессивному поведению. Полученные результаты представлены на рис. 7.1. Воспитание мышей A/J матерями из линии CFW существенно повышало показатель их агрессивности. Однако это изменение невелико по сравнению с уровнем межлинейных различий.
Воспитание мышей CFW матерями A/J не влияло на агрессивность потомства. Таким образом, различия между этими линиями в агрессивном поведении, по-видимому, обусловлены в основном наследственностью. Трансплантация яичников. Эксперимент с перекрестным воспитанием устраняет не поддающиеся контролю постнатальные влияния со стороны матери, однако пренатальные средовые условия при этом не изменяются. Метод трансплантации яичников позволяет отделить генетические эффекты от пренатальных материнских влияний. У животных инбредных линий происходит отторжение тканей, пересаженных им от животных других линий. Если, однако, скрестить две инбредные линии и получить гибридов первого поколения (Fi), то у этих гибридов трансплантаты яичников от обеих родительских линий не отторгаются. Скрещивая самок Fi, имеющих пересаженные «инбредные» яичники, с самцами той же инбредной линии, можно затем сравнить поведение мышей двух разных линий, развивавшихся либо у инбредных матерей, либо у матерей Fi. Используя этот метод, Де-Фриз и др. (DeFries et al., 1967) нашли, что в детерминации различий поведения в открытом поле главную роль играют генетические факторы, в то время как вес тела больше зависит от материнского влияния. Эксперименты с отбором Работы по искусственному отбору отличаются от попыток создать инбредные линии тем, что животных для скрещивания отбирают не на основе генотипа, а по фенотипу. Фенотипический признак – это любой доступный для измерения морфологический, физиологический или поведенческий признак организма. Вес тела, интенсивность обмена, активность в открытом поле – все это фенотипические признаки. Генотипом называют весь генетический материал организма или его часть. Для того чтобы начать эксперимент с искусственным отбором, нужно определить у группы самцов и самок из генетически гетерогенной популяции какой-либо признак. Самцов с высокими величинами данного признака скрещивают с такими же самками, а самцов с низкими величинами – с самками, у которых этот признак тоже слабо выражен. Потомство от скрещиваний тестируют на степень проявления того же признака, и для следующего скрещивания пары снова подбирают по такому же принципу. Таким образом в последовательных поколениях получаются две линии – одна в результате отбора на высокие показатели данного признака, а другая – на низкие. Для контроля изменений в условиях среды ведут еще третью линию – путем случайного скрещивания. Если в определении изучаемого признака имеется генетический компонент, показатели «высокой» и «низкой» линии в последовательных поколениях расходятся. Показатели линии со случайным скрещиванием не должны обнаруживать систематических отклонений. Де-Фриз, Хегман и Халком (DeFries, Hegmann, Halcomb, 1974) использовали метод отбора для анализа поведения мышей в открытом поле. Взяв в качестве исходного материала относительно гетерогенную группу, они в течение 20 поколений вывели две линии с высоким значением признака, две линии – с низким и две контрольные линии. Результаты представлены на рис. 7.2. Можно видеть, что за эти 20 поколений показатели признака для различных линий разошлись – они увеличились у животных с высоким значением признака и снизились в линии с низким значением, а у контрольных линий изменились очень мало. Эти данные иллюстрируют важную роль генов в контроле активности животных в открытом поле. Сходные результаты были получены в селекционных экспериментах с широким спектром видов и фенотипиче-ских признаков, включая фототаксис (тенденцию приближаться к источнику света) у плодовой мушки (Hirsch, Boudreau, 1958), научение крыс в лабиринте (Тгуоп, 1940), реакции, связанные с импринтингом, у цыплят (Graves, Siegel, 1969) и агрессивность у мышей (Lagerspetz, 1969).
Межвидовые гибриды В большей части исследовании по генетике поведения используют особей одного вида, имеющих разные генотипы. Однако в лабораторных условиях можно скрещивать особей разных, но близких друг к другу видов. В поведении потомков от такого скрещивания часто обнаруживаются явные признаки генетических влияний. Превосходный пример такого подхода – работа Дилгера (Dilger, 1962), который изучал поведение, связанное с постройкой гнезда, у разных видов неразлучников. Особи одного вида, Aga-pornis personata, отрывают кусочки материала для гнезда и переносят их к гнезду в клюве. Представители другого вида, А. го-seicollis, не держат их в клюве, а засовывают под боковые перья. Исследованные Дилгером гибриды f[ обнаружили смешанный тип поведения, при котором птицы пытались засунуть материал для гнезда в перья, затем вынимали его, брали их в клюв, а потом все начиналось сначала. Таким образом, их поведение представляло собой малоэффективную смесь из двух типов целесообразного поведения родительских форм. Сходные промежуточные формы обнаруживались в демонстрационном поведении вьюрков (Hinde, 1956) и в характере звуковых сигналов сверчков (Bentley, Hoy, 1974).
Менделевское скрещивание Чтобы выяснить, один или несколько генов определяют интересующий нас поведенческий признак, используют те же методы, которые применял в своей работе монах-августинец Мендель – основоположник научной генетики. Мендель работал с горохом и заметил, что некоторые его линии не дают расщепления по определенным признакам Например, все потомство от гороха с гладкими семенами имело гладкие семена, тогда как все потомство растений с морщинистыми семенами имело морщинистые семена. Мендель начал свои опыты со скрещиваний между такими линиями, не дававшими расщепления. При этом обычно все потомство оказывалось сходным с одной из родительских форм. При скрещивании гладкосеменной формы с растениями, дающими морщинистые семена, все потомки имели гладкие семена. Когда фенотип потомства от такого скрещивания сходен с одним из родительских фенотипов, этот наследуемый фенотип называют доминантным признаком, а тот, который утрачивается, – рецессивным. Далее Мендель скрещивал между собой растения Fi, чтобы получить второе поколение потомства – Fz. Результат был всегда один и тот же в потомстве встречались оба родительских типа – и с гладкими, и с морщинистыми семенами. Растений с доминантным признаком – гладкими семенами – было в 3 раза больше, чем с рецессивным – морщинистыми семенами. При скрещивании между собой растения Fz, имевшие рецессивный фенотип, уже не давали расщепления, тогда как в потомстве форм с доминантным фенотипом его сохраняла только одна треть растений. Эти результаты можно объяснить действием генов – неделимых дискретных единиц наследственности, постулированных Менделем Следует вспомнить, что гены представлены парами альтернативных форм – аллелей. Разные аллели обозначают разными символами, доминантные – заглавными буквами, а рецессивные – строчными. Генотип исходных растений с гладкими семенами мы могли бы записать как ГГ, поскольку этот признак доминантен и в потомстве не происходит расщепления. Генотип растений с морщинистыми семенами будет гг. Во втором поколении одна четверть растений будет ГГ, половина – Гг и еще одна четверть – гг Из такой модели вытекают предсказания, полностью соответствующие данным Менделя. Наследуемость В генетике поведения большое значение придается вычислению наследуемости. Эта величина говорит о том, какую долю общей изменчивости данного признака (в популяции) можно отнести за счет различий в генотипе. Нам нужно будет сначала рассмотреть метод оценки изменчивости – изменение вариансы. Варианса служит показателем дисперсии (или изменчивости) выборки. Предположим, что у нас есть две группы животных по три особи в каждой; животные одной группы посетили соответственно 49, 50 и 51 квадрат открытого поля, а животные другой группы – 30, 50 и 70 квадратов. Средние для обеих групп между собой равны, однако группы различаются по вариабельности отдельных значений относительно средней: во второй группе изменчивость больше. Варианса позволяет оценивать эту изменчивость количественно. Варианса – это средняя величина квадратов отклонений от среднего значения признака. Для вычисления вариансы находят разности между каждой величиной и средней, возводят их в квадрат и находят среднюю от этих квадратов. Вариансы двух приведенных выборок равны соответственно 0, 7 и 266, 7. Следует вспомнить, что всех особей одной инбредной линии можно считать генетически идентичными. Таким образом, в инбредной линии отсутствует генетическая варианса, поэтому как vq, так и CGD должны быть равны нулю Это не означает, что на данный признак генотип не влияет, а значит лишь, что в данной группе животных нет той доли изменчивости, которая определялась бы различиями в генотипе Если бы мы могли выращивать группу генетически гетерогенных особей в совершенно одинаковых условиях, CGD приближалось бы к 1. Это не значит, что средовые факторы для развития данного признака несущественны, а означает лишь, что в данной группе и в данных условиях различия между особями не определяются средой. Если мы обозначим генетическую аддитивную вариансу как va, то истинную наследуемость, или наследуемость «в узком смысле» (/i2), можно будет представить следующей формулой: Различие между va и vg состоит в том, что аддитивная варианса va. – это только часть всей генетической вариансы vg. Часть общей генетической вариансы связана с действием доминантных генов, подавляющих проявление своих рецессивных аллелей, а часть связана с эпистазом, т. е. взаимодействием между генами разных локусов Поскольку эта часть генетической вариансы следующему поколению непосредственно не передается, при статистической обработке ее исключают, чтобы получить величины аддитивной генетической вариансы и истинной наследуемости. Величина h2 всегда должна быть равна или меньше величины CGD, поскольку Уд – только часть общей генетической вариансы Ve-«Наследуемость признака – один из самых важных популяцион-ных параметров, которые можно оценить методами количественной генетики» (DeFries, Hegmann, 1970, р. 27). Вычисление наследуемости Наследуемость или CGD можно вычислять многими способами. Для людей можно использовать результаты близнецового анализа, корреляции между родителями и детьми и между сибсами. В исследованиях, проводимых на животных, обычно используют метод отбора, менделевские скрещивания или диаллельное скрещивание (см. ниже), чтобы находить величины h2 или CGD. У Фуллера (Fuller, 1960) мы находим пример вычисления CGD на основе данных менделевского скрещивания при изучении исследовательской ^активности мышей, помещенных в лабиринт Полученные данные представлены в табл. 7 2. Здесь приводятся средние и вариансы величин исследовательской активности для двух инбредных линий, BR и А, и для их гибридов Fi и F2 Для оценки CGD нам нужно знать величины оценок vq и Ут. Поколение Fz содержит практически все генотипы и поэтому поможет нам получить оценку V^. Поскольку все особи в каждой из инбредных линий и в гибридном поколении Fi генетически идентичны друг другу (все животные Fi гетерозиготны по всем локусам, по которым различаются инбредные линии), генетической вариансы внутри каждой из этих групп не будет.
Таблица 7.2 Исследовательская активности у мышей линий C57BR и A/Jax и их гибридов (Fuller, 1960)
Каждая из получаемых величин вариансы будет оценкой ve- Наилучшей оценкой ve будет средняя для трех варианс:
Генетическую вариансу Vo можно найти, вычтя ve из V-s (vg^v-s – ve; 29, 60 – 12, 82=16, 78). Тогда
Таким образом, мы установили, что несколько больше половины вариансы исследовательской активности у этих групп связано с генетическими факторами. Сходные величины были получены при исследовании мышей линий C57BL и BALB/c (DeFries, Hegmann, 1970). Хотя этот метод дает хорошую аппроксимацию величины CGD, его часто не удается применить в исследованиях по генетике поведения. Дело в том, что варианса Fz нередко бывает не выше вариансы Fi или родительских линий – так называемый «эффект Трайона» (Hirsch, 1967; DeFries, Hegmann, 1970). Это, видимо, связано с тем, что признаки поведения обычно находятся под влиянием большого числа генов. Наследуемость или CGD определяли, используя множество линий, методов и поведенческих признаков. Фуллер и Уаймер (Fuller, Wimer, 1973) подвели итог некоторым результатам; оказалось, что эти величины варьируют в пределах от 0, 22 для агрессивности у кур (оценка с использованием метода отбора) до 0, 89 для половой активности морских свинок (по данным скрещивания инбредных линий). Гены и биохимия Биохимически ген можно рассматривать как участок молекулы ДНК, кодирующий синтез определенного полипептида Молекулы ДНК, находящиеся в клеточном ядре, передают свою специфичность РНК (рибонуклеиновой кислоте). РНК направляет биохимическую активность цитоплазмы клетки, что в конечном счете приводит к образованию структурных белков и активных ферментов (см. Thiessen, 1972, McClearn, DeFries, 1973). Ферменты – это катализаторы биохимических реакций, скорость которых они изменяют. Эти реакции влияют на скелет, мышцы, эндокринные железы, вегетативную, периферическую и центральную нервную систему и т. д. – на все системы тела. Очевидно, что должны существовать вполне определенные пути от генов к ферментам, влияющим на биохимию мозга и поведение К сожалению, эти пути еще не вполне ясны. Последнее звено этой цепи будет рассматриваться в главе 11. Не все гены одинаковым образом влияют на синтез РНК. Гены, непосредственно участвующие в построении РНК, называются структурными генами Согласно модели Жакоба и Моно, существуют еще две важные группы генов Операторы осуществляют «включение» и «выключение» структурных генов. Гены-регуляторы действуют, усиливая или ослабляя активность операторов Таким образом, существует сложное взаимодействие между стимулирующим и тормозящим действием одних генов на другие. Их активность чувствительна к биохимическим факторам организма. Некоторые гены, по-видимому, включаются или выключаются на определенных стадиях развития или в определенных условиях При таком представлении о работе генов слова о том, что гены и условия среды непрерывно взаимодействуют, образуя единую динамическую систему, перестают быть риторикой или чистым теоретизированием: все это оказывается неизбежным результате сложных биохимических реакций. Глава 8 ОНТОГЕНЕЗ ПОВЕДЕНИЯ Изучение онтогенеза поведения включает анализ изменений, происходящих до вылупления из яйца или рождения, во время вылупления или рождения и в течение всего последующего развития. Онтогенез поведения в эмбриональный период В большинстве исследований объектом служили здесь домашние птицы Причина этого понятна. В скорлупе развивающегося яйца легко прорезать окошко, и если оно сделано аккуратно, можно непосредственно наблюдать ход развития поведения. Подобные манипуляции у живородящих млекопитающих значительно более трудны Подробно описан ход развития стереотипных двигательных актов у куриного эмбриона (Hamburger, 1963, 1971; Oppenheim, 1974) Хамбургер различает подвижность трех типов: Категория А – спонтанная подвижность Это результаты автономных (самоуправляемых) нервных разрядов, которые могут возникать в спокойных, неизменяющихся условиях среды (при отсутствии сенсорных стимулов) Категория В – эндогенно стимулируемая подвижность Это движения, которые запускаются или вызываются гуморальными агентами (жидкостями тела) или иными изменениями во внутренней среде Категория С – вызванная подвижность Сюда относятся движения, возникающие в результате стимуляции сенсорных систем внешними факторами У куриного эмбриона спонтанная подвижность начинается примерно через 3, 5 – 4 дня после начала инкубации в виде небольших наклонов головы Периоды таких движений возникают с нерегулярными интервалами, их длительность со временем постепенно увеличивается, а интервалы между ними сокращаются. Пик этой активности приходится на 13 и день Движения в это время охватывают голову, туловище, конечности, пальцы, глазные яблоки.. веки и клюв и обычно выглядят бесцельными, отрывистыми и судорожными. Начиная примерно с 17-го дня появляется совершенно иной тиго активности, для которого характерны высокоинтегрированные движения Хотя они возникают несколько реже, чем более ранние изолированные движения, они в конечном счете создают ориентацию тела, удобную для вылупления, после чего следуют проклевывание и сам акт вылупления (примерно на 20-й день). В последние несколько дней перед вылуплением зародыш издает звуки Позднее сходная в общих чертах картина была описана у лабораторной крысы (Narayanan et al, 1971). Хамбургер (Hamburger, 1971, р. 63) считает, что «спонтанная активность у эмбрионов предшествует активности в ответ на стимулы» Он подкрепляет этот вывод пятью группами данных 1) двигательная система дифференцируется раньше, чем сенсорная; 2) поэтому многие зародыши производят движения раньше чем становятся чувствительными к раздражению; 3) вплоть до 17-го дня развития куриного эмбриона нельзя обнаружить никакого влияния внешних стимулов на его спонтанную активность; 4) эмбрионы всех животных осуществляют спонтанные движения с первого момента появления подвижности, но только у млекопитающих они с самого начала чувствительны к сенсорной стимуляции; 5) способность к автономным электрическим разрядам, вероятно лежащим в основе спонтанной подвижности, – это, по-видимому, первичное свойство любой нервной ткани Хотя сенсорные стимулы как будто бы не нужны для развития спонтанной подвижности, возможно, что и стимуляция, и ранняя спонтанная подвижность необходимы для развития более поздних форм поведения (Gottheb, 1971). Об онтогенезе подвижности, стимулируемой эндогенно (категория В), мало что известно Вызванные движения (категория С) начинаются у куриного эмбриона примерно на 7-й день инкубации. Эмбрион реагирует на легкое прикосновение маленькой кисточкой,. а также и на более сильное раздражение Вначале эти движения можно бывает вызвать только в области рта, но уже спустя 1 – 2 дня чувствительная область расширяется и охватывает всю поверхность тела Чувствительность к проприоцептивным стимулам. появляется примерно на 10-й день. У цыпленка развитие чувствительности начинается с тактильной модальности, затем последовательно включаются вестибулярная, слуховая и зрительная системы (Gottheb, 1968, 1971). Поразительно то, что в таком же порядке, видимо, идет развитие сенсорных систем и у самых различных млекопитающих. Готлиб»-Gottlieb, 1971) высказал предположение, что эта последовательность является общей для птиц и млекопитающих и, возможно, установилась еще у их древних предков-рептилий, если не раньше. Юн пришел к выводу (Gottlieb, 1968), что лишение сенсорного входа или его обогащение в отношении лишь одной модальности вряд ли изменит порядок развития сенсорных систем у цыпленка, но может повлиять на относительную скорость развития разных систем. Эмбрионы пекинской утки способны избирательно реагировать на материнский сигнал утки их собственного вида примерно за 5 дней до вылупления. Эта способность не зависит от предварительной возможности слышать свои собственные звуки или звуки их сибсов. Однако изменение частоты и времени нормальной звуковой стимуляции позволяет предсказуемым образом ускорять или замедлять изменения в реакциях эмбрионов на материнские сигналы (Gottlieb, 1971). Поведение только что вылупившихся и новорожденных животных Представители разных групп позвоночных сильно различаются по уровню развития их поведения к моменту рождения или вылупления из яйца. У некоторых видов молодые особи появляются на свет достаточно зрелыми (зрелорождающиеся) и уже вскоре способны осуществлять значительное число двигательных актов, включая локомоцию. Это типично для многих птиц, живущих стаями (таких, как утки и куры), для стадных копытных (овец, коз, американских лосей), для морских свинок. У других видов птенцы или детеныши вначале относительно беспомощны (незрелорождающиеся), и для развития локомоции и других форм поведения им требуется определенное время. К незрелорождающимся относятся многие виды певчих птиц, такие, как крапивники и кардиналы, морские птицы, живущие колониями (например, чайки), и многие млекопитающие, в том числе крысы, мыши, кошки и большинство приматов. В период около момента вылупления в поведении цыпленка происходят большие изменения (Balaban, Hill, 1969; Clemens, 1973). У только что вылупившегося цыпленка обнаруживается резкое повышение уровня бодрствования и усиление мышечного -тонуса. После вылупления дряблый губкообразный эмбрион за несколько минут превращается в активный молодой организм. Хотя большая часть исследований по развитию поведения была проведена на позвоночных животных, хорошо известно, что только что появившаяся на свет молодь многих беспозвоночных способна «. относительно сложным интегрированным движениям. Например, бабочки рода Pieris способны к полету менее через 2 часа после выведения из куколки. «Для способности к полету не имеют большого значения ни созревание, ни упражнение» (Petersen, Lund-gren, Wilson, 1956, р. 338). Для новорожденных млекопитающих наиболее характерны два типа поведения – сон и сосание. У многих видов новорожденные проводят во сне две трети всего времени и больше. Скорость, с которой детеныши сосут молоко, варьирует от вида к виду, но остается более или менее постоянной в пределах одного вида (Wolff, 1968). Импринтинг (запечатление) Поведенческие акты птиц в природе адресуются другим организмам не случайным образом – обычно они направлены на биологически адекватные мишени. В своих вылазках из гнезда в ранний период жизни птенцы выводковых птиц следуют за матерью. По достижении половой зрелости большинство птиц выбирает брачного партнера только среди особей своего вида. Вопрос о природе процессов, обеспечивающих такую избирательность поведения, привлекал большое внимание исследователей. История вопроса. Ученые нашли много ранних упоминаний о явлениях, подобных импринтингу (например, Kevan, 1976). Из ранних работ наиболее содержательным было исследование Сполдин-ra (Spalding, 1873), который среди других явлений заметил, что, как только цыплята приобретают способность ходить, они начинают следовать за любым движущимся объектом. Хейнрот расширил наблюдения подобных явлений, исследуя множество разных видов птиц. Интерес к импринтингу больше всего стимулировали работы Конрада Лоренца (Lorenz, 1937a). Этот автор описал многочисленные случаи, когда птицы не спаривались с особями своего вида из-за того, что они были лишены контакта с ними в ранний период жизни. Например, Лоренц выращивал гусей в изоляции от сородичей в течение первой педели жизни и отметил, что такие птицы предпочитают следовать за людьми, а не за другими гусями. Птицы ждали перед парадной дверью дома Лоренца и пытались следовать за людьми, как только те выходили из дома. Лоренц назвал это явление Pragung, что по-немецки означает «впечатывание», а на английский было переведено как imprinting (запечатление). Лоренц высказал предположение, что импринтинг отличается от истинного ассоциативного научения четырьмя особенностями: 1. Импринтинг приурочен к очень ограниченному периоду жизни птицы – «критическому периоду». 2. Однажды совершившись, процесс запечатления далее необратим. Если у птенца произошел импринтинг по отношению к птице другого вида, то позднейший контакт с особями своего вида уже не сможет устранить этот эффект раннего опыта. 3. Импринтинг – уникальный феномен в том отношении, что объект, на который будет направлено поведение, может быть определен задолго до первого осуществления самого поведения. Было, например, показано, что ранний импринтинг, происходящий задолго до наступления половой зрелости, влияет на выбор полового партнера. 4. Лоренц рассматривал импринтинг как форму «супериндивидуального условного рефлекса» в том смысле, что поведение при этом будет направлено не на определенную особь, которую видел птенец, а на целый класс организмов, членом которого была особь, послужившая стимул-объектом. Здесь следует отметить, что в дальнейшем многие исследователи подвергли сомнению, действительно ли все эти отличия столь существенны Начиная с 1937 года импринтинг зарекомендовал себя как увлекательная проблема и для психологов, и для зоологов, которые в попытках понять это явление потратили немало часов и исписали немало страниц. Поскольку в работах по половому импринтингу и но запечатлснию привязанности использовались несколько различные методы и получены разные результаты, мы рассмотрим эти работы раздельно Запечатление привязанности. Говоря об этом явлении, мы рассмотрим проблемы, связанные с механизмом реакции следования у выводковых птиц. Говоря проще, нас интересует, как молодая птица узнает, за кем она должна следовать.
Рис 8 4 Аппарат для лабораторных опытов по изучению импринтинга (Hess, 1959) Птенец попадает на круговую дорожку и может видеть модель, движущуюся с постоянной скоростью. Методы. Большая часть исследований по запечатлению привязанности была выполнена в лабораториях, например в лаборатории Экхарда Хесса (Hess, 1959). Основной аппарат Хесса изображен на рис. 8.4. Птенец вылупляется из яйца в темноте, и его содержат изолированно в темном ящике внутри инкубатора вплоть до самого момента опыта. Через автоматическую дверцу птица попадает в аппарат на круговую дорожку диаметром около 1, 5 м. В большинстве экспериментов по дорожке с определенной скоростью перемещали чучело селезня (рис. 8.4), зеленую коробочку или красный мяч. В некоторые объекты был вмонтирован динамик, так что они могли издавать звуки. Птенец оставался в аппарате определенное время – на период запечатления. Если соблюдены все условия, он начинает следовать за движущимся объектом. Из аппарата птенца извлекают через дверку в полу. Позже его снова помещают в аппарат на время тестирования. При этом птенцу обычно предъявляют на выбор два объекта: тот, который ему ранее показывали, и какой-нибудь новый. Обычно птенец оказывает предпочтение объекту, который уже экспонировался. С помощью описанного аппарата можно проделывать много различных манипуляций Рассмотрим результаты некоторых экспериментов.
Стимул-объекты. Мозг птенца не tabula rasa (чистая доска) в том смысле, который придавал этому выражению Джон Локк. Хотя в экспериментах с имприитингом можно успешно применять Представлен процент птиц, обнаруживающих наилучшие показатели запечатления, как функция возраста.при первом предъявлении стимул объекта. самые различные стимул-объекты, некоторые из них более эффективны, нежели другие (Bateson, 1971; Graves, 1973). Стимулы, более заметные для человеческого глаза, оказываются более эффективными и для импринтинга. Например, Клопфер (Klopfer, 1967) нашел, что запечатление бывает значительно сильнее, когда используется модель, разрисованная в полоску, по сравнению с моделью, выкрашенной белой краской. Оказалось, что птицы, следовавшие в период импринтинга за белой моделью, в период тестирования предпочитали ей полосатую. Таким образом, хотя вообще птицы предпочитают следовать за объектами, на которые у них произошел импринтинг, мощные зрительные раздражители могут перекрывать эффект раннего опыта. Чувствительный период. В представлениях Лоренца об имприн-тинге важнейшую роль играл «критический период». В целом экспериментальные данные подтверждали, что наиболее эффективное запечатление происходит во время че'''ко ограниченной фазы развития организма. На рис. 8 5 представлены результаты, полученные Хессом. У утят готовность к импринтингу достигает максимума в период от 13 до 16 часов жизни. Воздействие стимул-объектов в более ранние и более поздние сроки неэффективно. Под «критическим периодом» обычно понимают тот период онтогенеза, когда определенная система наиболее подвержена модификации. Вероятно, в такие периоды происходит наиболее быстрое созревание механизмов, необходимых для данной реакции. Концепция критического периода вызвала много споров. Фактически все исследователи согласны в том, что запечатление. например, не происходит одинаково хорошо на всех стадиях жизни. Однако некоторые авторы утверждают, что изменения в чувствительности происходят несколько более плавно, чем можно было бы ожидать, исходя из концепции критического периода или из данных, представленных на рис. 8 4. Кроме того, оказалось, что для ряда поведенческих систем можно изменять временные характеристики «критического периода» Например, на период чувствительности к импринтингу могут влиять условия выращивания (Bateson, 1971). В связи с этим многие исследователи стали называть этот период не «критическим», а «чувствительным», чтобы не создавать представления о внезапности и жесткой фиксирован-ности происходящих изменений. Этот менее спорный термин мы и будем использовать. Однако это не должно умалять значение концепции об ограниченном периоде максимальной готовности к импринтингу. Запечатленные объекты как подкрепляющие стимулы. Показано, что объекты, на которые произошел импринтинг, служат для молодых птиц эффективными подкрепляющими стимулами. Если предъявлять такой объект после какой-либо произвольно выбранной реакции, птицы могут обучиться этой реакции Например, Хофман и др. (Hoffman et al, 1966) нашли, что цыплята клюют рычаг, если эта реакция ведет к появлению перед ними запечатленного объекта – пластиковой молочной бутылки. Процессы, лежащие в основе импринтинга. Вопрос о природе механизмов, лежащих в основе импринтинга, привлекал большое внимание и вызывал много споров. Недостаток места не позволяет нам рассмотреть этот вопрос подробно. Ражецкий (Rajecki, 1973) приводит шесть разных точек зрения относительно механизмов импринтинга: это интерпретации с точки зрения контекстуальной, этологической, условно-рефлекторной, эпигенетической и информационной концепций, а также концепции нейронных цепей Обратимость. Так же как и чувствительный период, свойство необратимости играло важнейшую роль в концепции импринтинга, предложенной Лоренцем. Поскольку содержать большое число животных в течение долгого времени довольно дорого, обратимость запечатления привязанности была предметом лишь немногих длительных исследований. Видимо, она варьирует в зависимости от природы раздражителя и условий импринтинга. В большинстве случаев запечатление привязанности, изучаемое в лабораторных условиях, оказывается легко обратимым Например, Хесс (Hess, 1972) показывал маленьким утятам людей в течение 20 часов непрерывно. Утята следовали за людьми всюду Однако они стали следовать за живой уткой-кряквой, после того как ее показывали им в течение полутора часов в более позднем возрасте. «Чисто лабораторный импринтинг обратим», – заключает автор (Hess, 1972, р. 25). И наоборот, Хесс не смог изменить импринтинг, который произошел у утят на их собственную мать-утку, которую они видели в течение 16 часов сразу после вылуп-ления. Межвидовые различия. Следует отметить, что разные виды в сходных ситуациях реагируют по-разному. Хотя очевидно, что у домашних цыплят и у утят кряквы закономерности импринтинга могут быть различными, игнорирование таких различий было причиной некоторой путаницы в литературе по импринтингу. Роль слуховых стимулов. Уже давно известно, что комбинированное воздействие зрительных и слуховых раздражителей при импринтинге более эффективно, чем действие одних только зрительных стимулов. Готлиб (Gottheb, 1973) сообщил о серии экспериментов, направленных на выяснение относительной важности тех и других раздражителей. Он исследовал птиц, которые вылуплялись и далее выращивались в условиях, когда только слышали, но не видели своих сибсов и никогда не видели взрослых особей и не слышали издаваемых ими звуков. Маленькие утята трех видов с готовностью следовали за моделью, похожей на утку-мать соответствующего вида, издававшей такие же звуки, как эта утка. Однако подмена зрительных компонентов такого стимул-объекта зрительным образом, характерным для другого вида, на реакцию следования почти не влияла. Напротив, изменение звукового сигнала существенно ослабляло реакцию следования. Птицы, никогда не слышавшие материнского зова, избирательно реагировали на крики особей своего вида Например, птенцы пекинской утки вначале следовали за одной из двух идентичных моделей матери, а затем, когда звуковые сигналы, издаваемые «матерями», менялись местами, утята либо изменяли предпочтение, либо просто переставали следовать за первой моделью. Эти наблюдения позволяют предполагать, что в ранний период жизни утят слуховая избирательность может быть первичной детерминантой реакции следования. Маленькие утята «попадают в ситуацию импринтинга уже „настроенными" на определенные аудиометрические характеристики материнского призывного сигнала своего вида» (Gottlieb, 1973, р. 371). Хесс показал, что между матерью-уткой и эмбрионом непосредственно перед вылуплением протекают настоящие диалоги. Хесс предполагает также, что таким путем молодые птенцы могут усвоить определенные звуковые характеристики сигналов именно их матери и таким образом окажутся способными отличать ее сигнал от сигналов птиц других видов Половой импринтинг. Говоря о «половом импринтинге», мы имеем в виду процесс, в результате которою данное животное начинает адресовать свое половое поведение одним особям, а не другим Проще говоря, птица должна определить, с кем ей надлежит спариваться Часто предполагают, что половой импринтинг и запечатление привязанности происходят в результате одного и того же процесса в те же самые сроки. Однако дело, видимо, не всегда обстоит так. Иногда эти два процесса могут быть разделены. Методы. Методы исследования полового импринтинга отличаются от тех, которые используют в случае запечатления привязанности. ^Обычно длительность экспозиции объекта в исследованиях по половому импринтингу значительно больше. Исследуя внутривидовой половой импринтинг, птицу выращивают с особями того же вида, но отличающимися по окраске (например, белыми). Такие эксперименты проводились с разными видами птиц – с утками, голубями, вьюрками. При изучении межвидового импринтинга птиц выращивают с особями других видов. Такие работы были проделаны с различными представителями утиных, куриных и воробьиных. Следует отметить, что здесь можно использовать птиц как выводковых, так и птенцовых видов. Особую категорию составляют эксперименты, в которых объектом запечатления служит человек. Хотя некоторые из подобных данных можно считать всего лишь анекдотическими, в настоящее время половой импринтинг на человека у выращенных в неволе птиц описан у 25 видов (Immelmann, 1972). При изучении полового импринтинга пробу на предпочтение полового партнера проводят в зрелом возрасте. Некоторые исследователи для ускорения полового созревания вводили птицам гормоны, однако такие процедуры могут исказить нормальный ход процессов развития. Стимул-объекты. Успешность полового импринтинга сильно варьирует в зависимости от использованного объекта. В некоторых работах, например, импринтинг на человека оказывается несколько более слабым, чем на особей другого вида, более похожих на птиц данного вида и поведением, и внешностью. Исследования по половому импринтингу на неодушевленные предметы, кажется, почти не проводились. Межвидовые различия. У разных видов относительная роль врожденных перцептивных образов и научения в феномене импринтинга весьма различна (Lorenz, 1937a, р. 268). Способность к запечатлению может варьировать внутри вида в зависимости от пола. Чувствительные периоды. Чувствительные периоды для полового импринтинга, по-видимому, менее жестко ограничены во времени и часто приходятся на несколько более поздний период жизни, чем периоды для запечатления привязанности. В большинстве исследований, в которых был обнаружен сильно выраженный половой импринтинг, экспозиция была сравнительно долгой. Половой импринтинг может происходить в совершенно иной период, нежели запечатление привязанности. По данным Иммельмана (Immelman, 1972), у серого гуся чувствительный период начинается около 50-дневного возраста и кончается примерно к 140-му дню, а у крякв, видимо, начинается между 5 и 19 днями жизни и длится несколько недель. Клингхаммер (Klinghammer, 1967) отмечает, что у галок, зебровых амадин и дубоносов половой импринтинг происходит относительно рано в гнездовой период, а у снегирей, ворон и воронов окончательный выбор полового партнера определяется перед достижением половой зрелости. У горлиц и других голубей на выбор полового партнера можно влиять еще долго после того, как молодые птицы становятся независимыми от родителей. Обратимость. Хорошо известно, что птицы, у которых произошел импринтинг на особей другого вида, могут спариваться также и с партнерами своего вида, если тестирование производится каждый раз только с одним из этих видов. Если же птице предоставить выбор, она часто проявляет предпочтение, которое, видимо, вполне устойчиво. Например, Шейн (Schein, 1963) исследовал трех индюков, которые в течение первых 32 дней жизни росли в изоляции от своего вида, а в возрасте 5 лет тестировались на предпочтение полового партнера. Птицы «ухаживали» за человеком в присутствии людей и особями своего вида, когда находились только с ними. В ситуации выбора они ухаживали только за людьми. Хотя в зависимости от длительности контакта, природы стимул-объекта и изучаемого вида результаты бывают разными, половой импринтинг все же оказывается достаточно устойчивым. Импринтинг у птиц. Обобщение. Литературу по импринтингу следовало бы рассмотреть подробнее. Это процесс очень сложный. Некоторые авторы склонны проводить аналогию между имприн-тингом у птиц и явлениями научения в раннем возрасте у других видов, в особенности у детей. При проведении таких аналогий запечатление у птиц обычно рассматривается как процесс, протекающий в пределах жестких сроков критического периода в раннем возрасте, совершающийся практически мгновенно наподобие «впечатывания» и совершенно необратимый. На самом же деле в зависимости от вида птиц, от стимул-объектов, типа изучаемого импринтинга и многих других факторов при запечатлении обнаруживаются совершенно различные функциональные особенности. При этом можно продемонстрировать полную гамму явлений, начиная от отсутствия предпочтения запечатленного объекта до аномального полового импринтинга, эффект которого сохраняется всю жизнь. Как в случае почти всякого " хорошо изученного поведенческого феномена, детерминанты оказываются сложными. Поспешное распространение выводов на другие виды и другие ситуации было бы здесь неоправданным. Импринтинг у млекопитающих. Явления, очень сходные с запе-чатлением привязанности у птиц, продемонстрированы у некоторых зрелорождающихся млекопитающих. Например, Слакин (Sluckin, 1968) смог показать, что у морских свинок обнаруживается дифференциальное предпочтение кубика, окрашенного в белый и черный цвета, по сравнению с теннисным мячом в зависимости от того, какой из этих предметов предъявлялся им в раннем возрасте. Клопфер (Klopfer, 1971) исследовал другой феномен – «материнское запечатление». Козы-матери избирательно реагируют на своих собственных детенышей, предпочитая их детенышам других самок. Все 15 коз, у которых детенышей отобрали сразу после родов, отказывались принять козлят, когда их возвращали спустя 1 – 3 часа. Напротив, 14 из 15 матерей, которым в сходном опыте был предоставлен пятиминутный период контакта с детенышами, козлят приняли. По-видимому, материнское запечатление – процесс очень быстрый. Влияние раннего опыта у млекопитающих В последнюю четверть века мы были свидетелями настоящего взрыва исследований по влиянию раннего опыта на последующее поведение млекопитающих. Целая армия психологов, работающих в области экспериментальной и сравнительной психологии, а также психологии развития, произвела на свет буквально сотни оригинальных и обзорных статей на эту тему. Стимулом для этого служили интересные данные об импринтинге у птиц, некоторые приложения теории Фрейда, а также очевидная важность сведений о влиянии условий воспитания в раннем возрасте на поведение взрослого человека. В этих работах использовались самые различные виды животных и типы экспериментальных воздействий. Поскольку на нескольких страницах невозможно дать полный обзор таких исследований, мы попытаемся лишь дать о них общее представление. Мы рассмотрим данные о влиянии некоторых воздействий на животное в пренатальном периоде, обогащения среды в раннем возрасте и депривации в этот же период. Пренатальные воздействия. Чтобы исследовать влияние пре-натальных факторов, беременную самку обычно подвергают действию того или иного стресса. Во многих случаях это приводило к стойким изменениям в поведении потомства. Наиболее распространенная гипотеза относительно механизмов такого эффекта состоит в предположении, что агенты, вызывающие стресс, изменяют гормональную среду организма самки, а это влияет на плод таким образом, что последствия проявляются на более позднем этапе жизни. Томпсон (Thompson, 1957) воздействовал на группу самок сочетанием звонка и электрического удара. Они научились избегать удара, перебегая на безопасную половину челночной камеры при звуке звонка. Во время беременности самок три раза в день подвергали действию звонка, но не давали ни тока, ни возможности осуществить реакцию избегания. Для исключения возможных постнатальных влияний потомство воспитывалось другими самками. Оказалось, что пренатальный стресс матери снижает уровень активности и удлиняет латентный период ее начала в тесте с «открытым полем»; удлиняется и латентный период выхода из обычной клетки для побежки за пищей после 24-часового голодания. Эти изменения были истолкованы как результат повышенной «эмоциональности» потомства тех матерей, которые были подвергнуты стрессу. Как и во многих других случаях, гены и внешняя среда здесь взаимодействуют, формируя определенный результат эксперимента. Де Фриз, Вейр и Хегман (DeFries, Weir, Hegmann, 1967) показали, что влияние физического или химического стресса, которому подвергали беременных мышей-самок, на поведение их потомства в тесте открытого поля зависит от линии отца, линии матери и типа стрессорного воздействия. Уорд (Ward, 1972) нашел, что стресс матерей в пренатальный период снижает у крыс-самцов копулятивную активность и способствует проявлению у них реакции лордоза, характерной для самок. Раннее обогащение среды. Огромное большинство работ по влиянию раннего опыта у млекопитающих касалось последствий обогащения среды в раннем возрасте. В лабораториях грызунов чаще всего содержат в относительно небольших стерильных металлических клетках. Под «обогащением» среды мы понимаем введение каких-либо факторов, дополняющих или усложняющих -обычную среду в условиях лаборатории. Литература по этому вопросу очень сложна, так как почти всем опытам, в которых обнаруживается тот или иной эффект, можно противопоставить другие опыты, в которых сходное воздействие этого эффекта не дает или даже вызывает противоположное изменение. Это обусловлено несколькими причинами. Основная трудность связана с большим числом переменных, которые могут влиять на поведение в более позднем возрасте, и с тем, что в разных экспериментах было использовано значительное число различных параметров. В обзоре ранних исследований по этому вопросу Кинг (King, 1958) отметил, что получаемые результаты зависели от возраста в период экспериментального воздействия, длительности этого периода, характера и интенсивности воздействия, возраста при тестировании и типа теста. У разных видов животных (обычно лабораторных крыс и домовых мышей) часто обнаруживался разный характер влияний. Нас не удивит, что и в пределах одного вида существует множество взаимодействий между генотипом и средой (Henderson, 1968a). Все это означает, что здесь трудно делать широкие обобщения. Мы должны учитывать все существенные факторы. Как мы уже видели в главе 7 при рассмотрении наследственности и психологических законов, природа организмов составляет важную часть всех законов. Эти сложности следует рассматривать скорее как интереснейшую проблему для дальнейшего изучения, а не как помеху в работе (Henderson, 1968a). Рассмотрим несколько важнейших факторов и некоторые результаты исследований в этой области. Экспериментальные воздействия. Во многих экспериментах ранние воздействия кажутся очень слабыми Можно в первые несколько дней жизни просто ежедневно забирать детеныша у матери и помещать его на несколько минут в жестяную банку. В других опытах на детеныша можно или воздействовать током, или систематически брать в руки и «ласкать», или делать что-нибудь еще во время изоляции от матери (рентгеновское облучение, введение веществ, вызывающих судороги, и т. п.). Разнообразные раздражители можно предъявлять, не забирая детеныша от матери. Чаще всего изучали влияние температурного стресса, но применялись также слуховой и другие типы стрессов. После прекращения вскармливания молоком детенышей можно выращивать в самых разнообразных условиях различной сложности. Влияние обогащения среды в раннем возрасте. Было описано множество эффектов, вызываемых обогащением окружающей среды в раннем возрасте Мы рассмотрим пять наиболее часто наблюдаемых явлений: 1) ускорение роста и развития; 2) ослабление эмоциональной реактивности; 3) улучшение результатов в задаче с научением; 4) более адекватные реакции организма на стресс; 5) изменения в развитии нервной системы (см. Daly, 1973). Часто оказывалось, что взятие детенышей в руки, удары тока и другие процедуры, проводимые в раннем возрасте, вызывают увеличение веса детенышей к моменту прекращения материнского вскармливания (см, например, Denenberg, Karas, 1961; Levine, Otis, 1958). Однако во многих работах таких эффектов получено не было (см. Daly, 1973). Сообщалось, что взятие в руки (handling) в раннем возрасте или удары тока влияют на «эмоциональность». Об изменениях в эмоциональности часто судят по поведению животных в «открытом поле». Хотя интерпретации, связанные с понятием эмоциональности, очень спорны, остается верным то, что, по многочисленным данным, в результате обогащения ранней среды происходят изменения частоты дефекации в открытом поле, уровне активности в том же тесте или обоих показателей (см, например, Denenberg, Smith, 1963, Denenberg et al, 1962) Удары тока или взятие в руки в раннем возрасте приводят к сходным изменениям. Оба этих воздействия обычно либо повышают активность, либо снижают частоту дефекации, либо вызывают и тот и другой эффект. Было неоднократно показано, что обогащение среды улучшает обучение целому ряду задач, особенно связанных с избеганием (см, например, Levine, 1956) Причина этого не всегда понятна. Возможно, что влияние обогащения внешней среды в период материнского вскармливания на научение вторична по отношению к другим изменениям, таким, как, например, изменения эмоциональных реакций в аппарате для тестирования Ряд работ показал, что после взятия в руки в раннем возрасте у крыс наблюдаются более адекватные реакции на стресс по сравнению с контрольными животными. Например, при помещении в новую обстановку они реагируют умеренным выбросом кортикостероидов, хотя в ситуациях с физической угрозой у них возникает более интенсивная реакция надпочечников, чем у контрольных животных (Levine, Mullins, 1966). Воздействия в раннем возрасте могут влиять на биохимию и анатомию мозга Тэпп и Марковиц (Тарр, Markowitz, 1963) сравнивали мозг крыс, подвергавшихся стимуляции, и крыс, которых в первые десять дней жизни никаким воздействиям не подвергали. У животных, которых брали в руки, вес подкорковых отделов мозга был больше, а уровень ацетилхолина в этих отделах – ниже. У крыс, выращенных в усложненной среде, отмечалось изменение ряда мозговых показателей, в том числе увеличение веса коры, повышение активности ацетилхолинэстеразы, увеличение толщины коры и более сильное ветвление дендритов нервных клеток (Rosenzweig et al, 1968; Greenough, 1975). Влияние времени воздействия. Почти все, кто изучает эффекты раннего опыта, согласны в том, что большое влияние на результаты оказывает время, когда производилось данное воздействие Многие из эффектов взятия в руки проявляются после осуществления этой процедуры в первые дни жизни, но не проявляются, если такому воздействию животных подвергают позднее. Обогащение внешней среды в пре- и постнатальный периоды может по-разному влиять как на биохимию и анатомию мозга, так и на поведение Хотя ясно, что возраст – важная переменная и что первые 5 дней жизни – это у мышей и крыс период особенно высокой чувствительности ко многим воздействиям, некоторые эффекты были обнаружены и при стимуляции в более поздние сроки. Поэтому Дененберг (Denenberg, 1968) пришел к выводу, что для эффектов обогащения внешней среды в раннем возрасте истинного критического периода не существует. Можно определить лишь период максимальной чувствительности к качеству и количеству стимуляции, к зависимым от нее переменным и другим подобным факторам Механизмы, лежащие в основе эффектов обогащения среды. Относительно этих механизмов было выдвинуто множество предположений (Russel, 1971; Daly, 1973). Мы рассмотрим три гипотетических фактора: 1) изменения температуры, 2) изменения поведения матери и 3) стресс в раннем возрасте. Согласно «гипотермической» гипотезе, как ее формулировал Шефер (Schaefer, 1968), влияние раннего опыта обусловлено изменением температуры Когда животное вынимают из гнезда и берут в руки, температура его тела изменяется. Для более поздних эффектов это изменение температуры может быть более важным, чем прямое влияние на нервную систему. Сходные эффекты были получены, когда животных подвергали только холодовому стрессу (см. Schaefer, 1968). Шефер считает, что нервная система новорожденного детеныша не настолько развита, чтобы реагировать на какое-либо прямое воздействие ранней стимуляции, и это делает роль температуры весьма правдоподобной. Второе объяснение состоит в том, что эффект раннего обогащения среды – результат изменений в поведении матери. Взятие в руки детенышей вызывает изменение в материнском поведении самки, в частности усиленное вылизывание детенышей (Priestnall, 1973; Lee, Williams, 1974). Животные, подвергающиеся многим воздействиям, издают особенно интенсивные ультразвуки, которые привлекают повышенное внимание матери. Таким образом, изменения в поведении могут быть в большей мере обусловлены усиленным вниманием со стороны матери, чем прямым воздействием на детеныша (Richards, 1966; Barnett, Walker, 1974; Thoman, Levin, 1970). Третья точка зрения исходит из наблюдений, касающихся стресса. Мы уже отмечали, что животные, которых в раннем возрасте брали в руки, более «адекватно» реагируют на умеренный и сильный стресс, нежели контрольные особи. Регуляция уровня гормонов осуществляется, по-видимому, с помощью отрицательной обратной связи через гипоталамус и гипофиз (см. гл. 12). При повышении уровня гормонов активность центров, управляющих секрецией гормонов, снижается. Эта система отрицательных обратных связей, напоминающая термостат, регулирующий температуру воздуха в помещении, была названа «гормоностатом» (Le-vine, Mullins, 1966). Предполагается, что в результате повышения активности надпочечников, вызванного стимуляцией в раннем возрасте, какие-то характеристики гормоностата изменяются так, что у взрослого животного он действует более эффективно. Эти три гипотезы, а также другие объяснения эффектов раннего опыта не исключают друг друга. В определенных случаях могут действовать все эти факторы, играя различную относительную роль в зависимости от ситуации. Влияние, осуществляемое через мать. Мы уже видели, что воздействие на детенышей влияет на поведение матери и что изменения материнского поведения могут служить важным посредником в механизме многих эффектов от обогащения внешней среды в раннем возрасте. Следующий вопрос касается возможных изменений в поведении детенышей, связанных с теми воздействиями, которые претерпевали их матери, когда в свою очередь были детенышами. Дененберг и Уимби (Denenberg, Whimbey, 1963) показали, что потомство матерей, которых брали в раннем возрасте в руки, отличалось от потомства контрольных животных и по весу тела к концу материнского вскармливания, и по показателям «открытого поля». Дененберг и Розенберг (Denenberg, Rosenberg„ 1967) сделали еще один шаг дальше и показали, что такое воздействие на крыс-самок влияет на активность и вес тела при отъеме от матери у их внуков. Суммарный эффект определялся взаимодействием ряда переменных. Интерпретация. Все согласны в том, что обогащение внешней среды в раннем возрасте глубоко влияет на поведение взрослого животного. Относительно природы переменных, вызывающих такие изменения, тоже нет больших разногласий. Однако эти переменные, по-видимому, взаимодействуют в различных условиях по-разному, и это приводит к невероятному разнообразию конкретных результатов и порождает много споров относительно их. интерпретации. Нужно сделать еще одно предостережение. Обычно принято Считать влияние раннего опыта «благоприятным», а условия содержания в лаборатории – «обедненными». Как подчеркнул Дэй-ли (Daly, 1973), такие обобщения следует делать с большой осторожностью. Мы не можем уверенно сопоставлять условия большинства лабораторных тестов с факторами, существенными для-выживания и успешного размножения животных в их природной среде. Поэтому трудно что-либо заключить о том, что окажется благоприятным для них в естественных условиях. Такая же осторожность нужна при оценке условий содержания животных в лаборатории. Большинство грызунов живет в норах. Эти норы устроены так, что колебания температуры в обычных лабораторных условиях могут быть значительно большими, чем в норе. Весьма вероятно, что лабораторная клетка создает для детенышей-сосунков более стимулирующее сенсорное окружение, чем нора. Дэйли (Daly, 1973, р. 435) предполагает даже, что «условия содержания в лаборатории оказываются для очень маленьких детенышей чрезмерно стимулирующими». Обеднение среды, (условия депривации). Очевидным дополнением к экспериментам с обогащением внешней среды служат опыты с ее обеднением. В типичном случае детенышей выращивают & условиях, когда они либо изолированы от особей своего вида, либо лишены каких-либо раздражителей. Многие из таких исследований были специально посвящены изучению онтогенеза перцептивных систем и будут обсуждаться в главе 10; здесь же мы рассмотрим влияние обедненной среды на эмоциональность и на общественное и репродуктивное поведение. Собаки. Томпсон и Хероп (Thompson, Heron, 1954) нашли, что шотландские терьеры, выращенные в условиях с рядом ограничений, решали задачи хуже, чем собаки, выросшие в домашних условиях. Мельцак (Melzak, 1968) суммировал результаты нескольких исследований, в которых собак выращивали в условиях с резко ограниченным сенсорным входом. После снятия этих ограничений такие собаки обнаруживали: 1) чрезвычайно высокий уровень активности, 2) аномальное восприятие зрительных, слуховых, обонятельных и болевых стимулов и 3) трудности в торможении неадекватных реакций. Например, они с трудом научались тормозить реакцию приближения к зажженной спичке. Как показал Фуллер (Fuller, 1967), изоляция в раннем возрасте сходным образом влияет на поведение животных во взрослом состоянии. Он предложил три альтернативных объяснения этих эффектов. Первое было связано с представлением о том, что внешние стимулы необходимы для развития нормальной организации нервной системы; тогда аномальное поведение – это результат недостаточного созревания нервной системы, для которого нужны индуцирующие сенсорные воздействия. Второе объяснение может состоять в том, что нервная система при рождении уже организована, но из-за отсутствия сенсорного входа претерпевает своего рода дегенерацию. В качестве третьей альтернативы Фуллер предположил, что, когда содержавшееся в изоляции животное впервые попадает в нормальную обстановку, воздействие неожиданного потока сенсорной информации может само по себе вызывать аномальное поведение. С этой точки зрения влияние сенсорной депривации связано не столько с нею самой, сколько с резким переходом от обедненной среды к нормальной. Фуллер показал, что ежедневного 10-минутного пребывания собаки в нормальных условиях в течение двух недель оказалось достаточно, чтобы устранить многие из эффектов депривации. Особое обращение с животными и применение транквилизаторов во время выхода из условий депривации оказывали действие, противоположное эффекту депривации. Макаки-резусы. После работ по импринтингу наиболее известными экспериментами с изменением раннего опыта, по-видимому, являются работы Гарри Ф. Харлоу и его сотрудников на макаках-резусах (например, Harlow, 1962, 1971; Harlow, Harlow, Suomi, 1971, Mason, 1960). Эти исследователи выращивали обезьян в условиях разной степени депривации. Многие были выращены в полной изоляции от других особей. В таких опытах использовали модельных «матерей», покрытых плюшем. Такие «матери» были необходимы многим детенышам как предмет, к которому можно было прижиматься и проявлять другие реакции, выражающие привязанность и в норме адресованные матери. Когда, однако, выращенные в условиях депривации обезьяны достигали зрелости, у них обнаруживались сильные нарушения репродуктивного поведения. Некоторые обезьяны не только отказывались спариваться, но и в более раннем возрасте обнаруживали склонность к аномальным манипуляциям с ротовой полостью, сжимали руками собственное тело, раскачивались, обнаруживали «апатию и безразличие к внешней стимуляции» (Harlow, Harlow, Suomi, 1971, р. 543). С большим трудом удалось добиться спаривания нескольких самок. Оказалось, что эти «матери без матерей» совершенно не способны выполнять свои функции: они игнорировали детенышей, жевали их ладони и ступни, бросали их лицом на пол. Если выросшую в изоляции обезьяну помещали с нормальными сверстниками, она навлекала на себя сильные проявления агрессии. Однако контакт с обезьянами более молодого возраста оказывал «лечебное» действие. Грызуны. В опытах с ранней депривацией, проводимых на грызунах, изучали главным образом последующее репродуктивное поведение. Хотя ранние исследования как будто говорили о другом, нам теперь ясно, что у выращенных в изоляции крыс и морских свинок обнаруживаются нарушения в акте спаривания (Valen-stein et al., 1955; Gerall, Ward, Gerall, 1967; Gruendel, Arnold, 1969). Обычно у них наблюдаются аномальные движения и отсутствие правильно ориентированной садки, так что копуляция оказывается неэффективной. Хотя у выращенных в изоляции самок не было видимых дефектов в материнском поведении, у их потомства обнаруживалась повышенная смертность, а также задержка в сроках открывания глаз по сравнению с потомством нормальных самок (Thoman, Arnold, 1968). КРАТКИЕ ВЫВОДЫ Онтогенетические изменения поведения можно проследить в течение всей жизни животного. В эмбриональном развитии появлению вызванных движений предшествует спонтанная подвижность. Постнатальное созревание связано с более медленными и постепенными изменениями в разнообразных формах поведения, включая сон, рефлексы, двигательную активность и общественное поведение. Импринтинг в форме «запечатления привязанности» могут вызывать весьма различные стимул-объекты, и он наиболее эффективен в довольно короткий чувствительный период. Эффект запечатления привязанности, по-видимому, обратим и может быть снят в результате предпочтения определенных звуковых сигналов, которое, видимо, бывает врожденным. Для полового импринтинга чувствительный период более длителен и наступает позднее, но этот вид импринтинга во многих случаях не так легко обратим. Обогащение среды и депривация в раннем возрасте сильно сказываются на поведении животных в более позднем возрасте. Специфические эффекты, получаемые при ранних средовых воздействиях, варьируют в зависимости от таких переменных, как возраст во время воздействия и во время тестирования, характер воздействия и природа зависимой переменной.
Глава 4 ФОРМЫ ИНДИВИДУАЛЬНОГО ПОВЕДЕНИЯ В этой главе мы рассмотрим некоторые формы поведения, проявляемые отдельными особями в процессе адаптации к окружающей их среде. Это будут действия, с помощью которых животное получает пищу, воду, кислород и другие источники энергии; поддерживает чистоту своего тела; взаимодействует с окружающей средой, а также определенным образом организует свою активность. Все эти формы поведения имеют решающее значение для выживания. Они связаны с рядом конкретных функций, к которым относятся: 1) локомоция; 2) питание и дыхание; 3) терморегуляция; 4) поиски убежища; 5) избегание хищников; ч 6) сон; 7) поддержание чистоты тела; 8) выделение; 9) исследовательская активность; 10) игра; 11) использование орудий; 12) биологические ритмы.
Репродуктивному и общественному поведению будут посвящены главы 5 и 6. Здесь нужно сделать три замечания. Во-первых, при распределении материала по трем главам этой части и по разделам внутри глав мы исходили из соображений удобства и ясности изложения. Многие разграничения несколько произвольны, а некоторые из них в лучшем случае неоднозначны. Например, поиски укрытия могут иметь важное значение для того, чтобы избежать хищников; исследовательское поведение может составлять часть игры; потребление пищи влияет на терморегуляцию. Практически все эти типы поведения (например, питание, поиски укрытия, сон) иногда могут носить ярко выраженный общественный характер. Даже акты выделения могут иметь большое социальное значение, как, скажем, использование мочи для того, чтобы пометить занятую территорию. Таким образом, функциональное подразделение проводится здесь исключительно для удобства изложения, и его не следует принимать как безусловное. Во-вторых, следует помнить, что «животное – это приспособленный организм, а не просто совокупность специализированных адаптации» (Bunnell, 1973, р. 79). Способы приспособления животного к различным аспектам окружающей его среды переплетаются и взаимодействуют, образуя единое функциональное целое. Например, способ питания данного организма влияет на характер передвижения, на способ избегания врагов и на особенности сна и в свою очередь подвержен влиянию каждого из этих видов поведения. Решающий критерий – выживание, а размножение служит «нижним пределом». Действие отбора направлено на создание целостных организмов, способных функционировать в среде, где они обитают, а не на полное совершенство в какой-либо отдельной области. В-третьих, совершенно очевидно, что исчерпывающим образом изложить все эти вещи в одной главе невозможно. Мы попытаемся лишь дать некоторое представление о разнообразии форм поведения в пределах данной его категории, а также приведем по нескольку интересных примеров. Локомоция Локомоция имеет большое значение для большинства видов поведения, . так как она позволяет животному перемещаться в пространстве. Типы локомоции Существует пять основных типов локомоции: 1) движение при помощи жгутиков или ресничек; 2) амебоидное движение, осуществляемое путем изменения формы тела; 3) волнообразное движение; 4) реактивное движение и 5) движение при помощи конечностей (Maier, 1970). При движении первого типа животное передвигается в результате биения либо одного жгутика, либо группы волосовидных ресничек; такое движение широко распространено у простейших У беспозвоночных часто встречается также перемещение путем изменения формы тела, например путем вытягивания псевдоподий у амебы. Движение за счет волнообразных сокращений, пробегающих по телу, особенно характерно для змей, водных млекопитающих и рыб. При реактивном движении вода с силой выталкивается из организма в окружающую среду; этот тип локомоции используется у ряда беспозвоночных, таких, как медузы и кальмары Передвижение с помощью конечностей, будь то ноги, крылья или плавники, характерно для большинства позвоночных и для некоторых беспозвоночных. Передвижение при помощи конечностей Локомоция в результате движения конечностей может осуществляться в воде, на деревьях, в воздухе, под землей или на поверхности земли В воде. Плавание с использованием конечностей свойственно моржам и многим другим позвоночным. Разнообразные млекопитающие, живущие в основном на суше, в том числе многие грызуны, обезьяны и хищники, в случае необходимости достаточно хорошо плавают. На деревьях. Для большинства видов, обитающих на деревьях, характерно наличие цепких когтей, а также хорошо развитые зрительная система и вестибулярный аппарат. Многие виды лазают по стволам либо лазают или бегают по веткам Некоторые обезьяны способны к брахиации – передвижению при помощи передних конечностей, поочередно закидываемых вперед, тогда как тело висит в воздухе В воздухе. Хотя представители ряда различных групп, например летучие рыбы и летяги, способны к планированию, главной формой передвижения в воздухе служит истинный полет, наблюдаемый у насекомых, птиц и летучих мышей. У птиц наиболее распространенный тип полета – это машущий полет, при котором птица ритмично поднимает и опускает крылья. При этом внутренние части крыльев обеспечивают главным образом создание подъемной силы, преодолевающей силу тяжести, а наружные части создают пропеллирующий эффект, толкая птицу вперед (Hein-roth, Heinroth, 1958). Под землей. Роющими или полуроющими называют виды, которые всю свою жизнь или большую ее часть проводят под землей. У таких животных обычно имеются многочисленные адаптации (малая величина глаз и ушных раковин, менее густая шерсть и др), уменьшающие трение при передвижении под землей. На поверхности земли. Существует много разных типов передвижения животных по земле У ряда различных млекопитающих наблюдается хождение на двух ногах, наиболее характерное для некоторых кенгуру и грызунов, а также для человека Такие животные, как кенгуру и кенгуровые крысы, передвигаются прыжками Формы локомоции у большинства видов, передвигающихся на четырех конечностях, довольно стереотипны. При ходьбе конечности переставляются в таком порядке, чтобы всегда сохранялась опора в виде треугольника, создаваемого тремя конечностями. При большей скорости движения, например при рыси и галопе, устойчивость снижается (Gray, 1953). Гну и львы способны бежать со скоростью до 80 км/ч, а гепарды – даже до 110 – 120 км/ч (Bourliere, 1964). ПИТАНИЕ И ДЫХАНИЕ Потребление пищи, питье воды и дыхание имеют решающее значение для выживания самых разнообразных животных. Потребление пищи Стратегии питания. Животных можно разделить по их главной стратегии питания на пять основных групп, фильтрующие воду, паразиты, растительноядные, плотоядные и всеядные (Maier, Maier, 1970). Все фильтрующие организмы живут в воде и добывают себе пищу, отцеживая ее из воды Движение воды относительно тела животного достигается либо путем того или иного нагнетающего действия, либо за счет плавания Паразиты, как правило, живут целиком за счет организма-хозяина; некоторые из них (например, лентецы) живут в течение долгого времени, не убивая своего хозяина, а другие довольно быстро вызывают его гибель. Растительноядные питаются растениями, их корой, листьями, семенами, плодами и корнями. Живущий в Австралии коала высо-коспециализирован в отношении пищи и питается исключительно листьями эвкалипта У жвачных имеется четырехкамерный желудок, обеспечивающий более полное переваривание пищи. Плотоядные питаются главным образом другими животными Как среди беспозвоночных, так и среди позвоночных можно найти виды с самыми разнообразными типами питания. Многие животные всеядны, они поедают всевозможные виды пищи. У таких животных стратегии питания наименее специализированы Например, бродячие муравьи уничтожают почти все, что попадается на их пути в тропическом лесу Весьма разнообразные формы всеядности можно встретить у грызунов (Landry, 1970) Даже шимпанзе, по-видимому, всеядны, причем для них характерен удивительно высокий уровень организации охоты (Teleki, 1973а, Ь). Формы поведения, связанные с питанием. Из таких форм поведения мы рассмотрим хищничество, запасание пищи, разного рода манипуляции с нею и регулирование ее потребления. Хищничество. Некоторые виды, например пауки, строящие паутину, богомолы и муравьиные львы, пассивно поджидают добычу, тогда как большинство видов активно охотится. Пойманную добычу хищник хватает, душит, сжимает, отравляет, обездвиживает, оглушает или поедает живьем. Диапазон приспособлений к хищничеству поистине велик. Последовательность действий хищного млекопитающего можно разбить на приближение к жертве, ее умерщвление и поедание (Ewer, 1968). Домашние кошки подкрадываются к своей жертве, прижимаясь к земле, следя за каждым ее движением и используя короткие перебежки, чтобы все ближе и ближе подойти к ней, а затем совершить нападение. В отличие от этого гепарды приближаются к жертве, преследуя ее в открытую, и используют свою исключительную скорость бега лишь в том случае, когда жертва обращается в бегство. Млекопитающие часто убивают свою жертву, прокусывая ей затылок. Совершаемые при этом действия различаются в зависимости от вида хищника и от обстоятельств. Иногда хищник сначала схватывает жертву в лапы или прижимает ее к земле. Для многих видов, в частности для собачьих, характерно ритуализо-ванное «умерщвление встряской»: схватив жертву в зубы, хищник размахивает ею из стороны в сторону (Eisenberg, Leyhausen, 1972). Большинство хищников сначала поедает голову жертвы, а затем постепенно съедает ее всю в направлении от головы к заднему концу тела. Как показал Лейхаузен, ориентация жертвы обусловливается прежде всего осязательными стимулами – расположением шерсти или оперения. Поедание крупной добычи часто начинается с брюха или паховой области (Ewer, 1968). Насытившись, хищник обычно прекращает поиски пищи и охоту. Лишь в редких случаях он убивает больше того, что способен съесть;, это может произойти в условиях, когда плотность популяции жертвы очень велика и поэтому усилия, затрачиваемые на ее поиски, минимальны (Kruuk, 1972). Запасание пищи. Поскольку количество пищи в разное время года неодинаково, многие животные, особенно грызуны, делают запасы. Существуют две главные формы запасов: сконцентрированные в специальных «кладовых» и рассеянные по разным местам отдельными кучками (Ewer, 1968). Кладовые обычно устраиваются в самой норе, часто в специальной камере, а при рассеянном хранении запасов они могут быть разбросаны по значительно более обширной площади и находиться довольно далеко от дома. Встречаются и промежуточные стратегии. В норах прерийных полевок – небольших грызунов, обитающих в полях, – находили запасы хорошо сохранившихся семян объемом до 9 – 10 литров (Jameson, 1947). У многих видов, запасающих корм, имеются обширные защечные мешки, помогающие животным переносить корм к месту его хранения. Запасание корма у грызунов можно вызвать в лабораторных условиях; изучалось влияние, которое оказывают на эту форму поведения лишение пищи, индивидуальный опыт в ранний период жизни, различные физиологические воздействия и другие подобные переменные. Манипуляции с пищей. Характер манипуляций, производимых с пищей до ее потребления, сильно варьирует у разных видов и зависит от типа пищи. Так, например, калан ложится в воде на спину, кладет себе на грудь плоский камень и вскрывает раковины моллюсков, ударяя ими об этот камень. Другие животные используют для разного рода манипуляций передние лапы и пасть. Для развития многих типов манипуляций с пищей, по-видимому, достаточно лишь минимального опыта..Раскалывая орехи, белки прогрызают в скорлупе одну или две бороздки, расположенные определенным образом. Эйбл-Эйбесфельдт (Eibl-Eibes-feldt, 1963) вырастил в неволе белок, которым не приходилось прежде раскалывать орехи, и когда животные впервые приступили к этому, разгрызающие движения в общем не отличались от обычных, хотя были довольно плохо ориентированы. Регуляция потребления пищи. Животные не кормятся непрерывно. По мере того как проходит все больше времени после кормежки, особи многих видов становятся более активными и начинают особенно живо реагировать на стимулы, ассоциированные с пищей. Потребление пищи вовремя прекращается; у большинства животных проблема ожирения возникает редко. У млекопитающих потребление пищи регулируется, по-видимому, в результате взаимодействия двух ядер гипоталамуса – латерального и вентромедиального (Keesey, Powley, 1975). В отличие от этого у падальных мух потребление пищи регулируется в основном периферически (т. е. меньше зависит от центральной нервной системы): его инициируют периферические рецепторы, а тормозят импульсы, поступающие от растянутой передней кишки (Dethier, 1967). Потребление воды Вода совершенно необходима для самых разнообразных жизненных процессов, в том числе для пищеварения, выделения и терморегуляции. Многие животные должны активно поглощать воду как таковую, тогда как другим достаточно воды, извлекаемой в процессе метаболизма из пищевых веществ. Разные животные используют при питье различные движения. Собаки и волки принимают стереотипную позу и лакают воду языком, тогда как корова всасывает воду ртом, а язык играет у нее лишь пассивную роль. У большинства грызунов наблюдается довольно стереотипное слизывание воды. Слоны, конечно, используют хобот, чтобы насосать воду и перенести ее в рот (Bunnell, 1973). Дыхание Потребление кислорода путем дыхания – явление столь универсальное, что его нередко упускают из виду. Практически все животные обладают тем или иным механизмом, при помощи которого свежий воздух поступает в организм, а использованный выводится наружу. Дыхание становится проблемой первостепенной важности у водных млекопитающих. Дыхательные системы у них обладают удивительными приспособлениями, позволяющими этим животным нырять на большие глубины и оставаться под водой дольше, чем это могут другие млекопитающие. По имеющимся данным, ондатра и морской слон способны оставаться под водой в течение 12 минут, тогда как кит-бутылконос может погружаться на 120 минут (Bourliere, 1964). ТЕРМОРЕГУЛЯЦИЯ Животные способны существовать только в определенном диапазоне температур. Регуляция температуры тела – важнейший аспект приспособления к среде, особенно в суровых условиях пустыни или Арктики. Птицы и млекопитающие – гомойотермные животные; у них имеются физиологические механизмы, поддерживающие температуру тела на относительно постоянном уровне, несмотря на резкие колебания температуры среды. В отличие от этого беспозвоночные, рыбы, амфибии и рептилии пойкилотерм-ны. – температура тела у них колеблется в зависимости от внешней температуры. Многие формы поведения по крайней мере частично связаны с терморегуляцией. Для многих видов характерны сезонные миграции, назначение которых в том, чтобы сводить к минимуму воздействие неблагоприятных температур в определенное время года. Более кратковременная регуляция температуры тела достигается путем значительных ежедневных перемещений из затененных участков в освещенные или из норы наружу и обратно. В лабораторных условиях животное может научиться выполнять произвольные действия, если изменения температуры используются в качестве подкрепления (см., например, Wright, Meyer, 1969). Многие виды переживают суровые зимы, впадая в зимнюю спячку. При этом животное прекращает всякую активность, а температура тела, частота сердечных сокращений и уровень метаболизма снижаются у него до минимума. Некоторые виды лягушек, крокодилов и рыб реагируют аналогичным прекращением активности и на крайне высокие температуры – это так называемая летняя спячка. ПОИСКИ УБЕЖИЩ Большинство видов предпринимает поиски каких-либо убежищ, чтобы укрыться от резких колебаний температуры, осадков и хищников. Иногда животное просто забирается в пещеру, расселину или на дерево, что не приводит к сколько-нибудь существенному изменению среды. Однако во многих других случаях оно строит весьма сложные гнезда или норы, что влечет за собой значительные изменения внешних условий. Постройки бывают относительно постоянными, как, например, плотины бобров, или временными, как спальные гнезда шимпанзе, в которых они обычно проводят лишь одну ночь. У многих видов устройство гнезда тесно связано с размножением: когда близится время появления потомства, они приступают к постройке гнезда или значительно расширяют уже имеющееся гнездо. Среди беспозвоночных постройка убежищ наиболее ярко выражена у насекомых. У таких видов, как одиночные осы, каждая отдельная самка роет норку и запасает в ней корм. Однако у многих других видов гнезда представляют собой весьма сложные постройки, и в них обитают целые сообщества. Примером служат высокие сооружения термитов и гнезда пчел. Структура сообщества у разных видов пчел различна, но в них непременно имеется одна матка и много рабочих особей. У медоносной пчелы отличительная черта деятельности рабочих особей по постройке и содержанию гнезда – функциональная специализация. Сооружением сотов, кормлением личинок, уборкой ячеек, приготовлением меда, охраной входа и сбором пыльцы и нектара занимаются разные особи. При этом функции каждой отдельной рабочей пчелы изменяются в течение ее жизни: она начинает с чистки ячеек и заканчивает сбором пыльцы и нектара (Lindauer, 1961). Выбор места для нового гнезда у медоносной пчелы – процесс весьма интересный. В конце весны матка и около половины рабочих особей оставляют старое место дочерям матки и образуют рой на небольшом расстоянии от него. Пчелы остаются в этом рое до тех пор, пока не будет выбрано новое место. Пчелы-разведчицы вылетают из роя для обследования многочисленных мест, потенциально пригодных для устройства гнезда. Возвращаясь в рой, они исполняют «танец», содержащий указания о том, где находятся эти места (см. гл. 5). Интенсивность танца варьирует в зависимости от качества данного места. Особое значение придается, по-видимому, его величине и защитным возможностям. Танцующие пчелы вербуют новых разведчиц. На основании интенсивности танцев и реакции новых разведчиц рой «принимает решение»: в конце концов преобладающая часть разведчиц указывает при помощи танца одно определенное место, и тогда рой снимается и направляется туда (Lindauer, 1961). У грызунов поведение, связанное с поиском или постройкой убежища, принимает весьма разнообразные формы. Бобры строят однокамерные норы, или хатки, в которых обитают пара взрослых особей и два их последних выводка. Древесные крысы собирают множество прутиков или веточек, из которых строят обширные шалаши. Подробно изучена постройка гнезд лабораторными крысами и домовыми мышами. Оба вида устраивают гнезда в форме кубков или чаш, используя вату, бумагу, тряпки и другие подобные материалы; иногда гнезда снабжаются крышей. Шимпанзе, орангутаны и гориллы строят себе «спальные» гнезда на деревьях. ИЗБЕГАНИЕ ХИЩНИКОВ Поскольку большинство видов служит добычей по меньшей мере для нескольких других видов, избегание хищников имеет важное значение для выживания и размножения. Главные способы защиты от хищников – это укрывание от них, предупреждение особей своего вида, наличие предостерегающих признаков, бегство и активное сопротивление. Укрывание Многие животные скрываются от хищников в убежищах – норах, щелях и шалашах. Кроме того, укрытию от хищника может способствовать внешний вид самого животного. Покровительственная окраска, благодаря которой животное сливается с фоном, встречается у представителей почти всех таксономических групп. Много особенно ярких примеров можно найти у насекомых, среди которых есть формы, сходные с листьями, веточками или даже с птичьим пометом. Часто покровительственная окраска сочетается с особым поведением: животное располагается по отношению к окружающему определенным образом, обычно сохраняя неподвижность.
|
Последнее изменение этой страницы: 2019-04-09; Просмотров: 756; Нарушение авторского права страницы

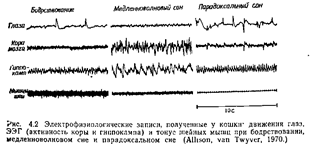
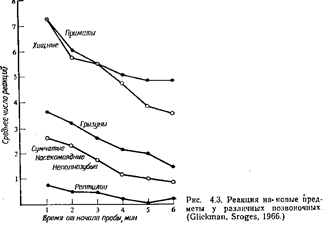
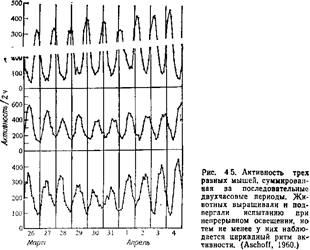
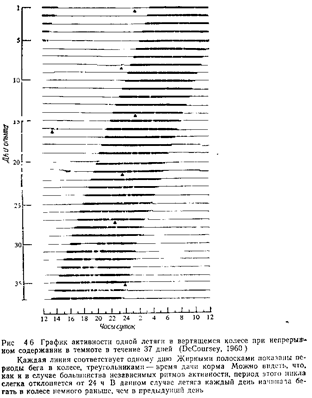

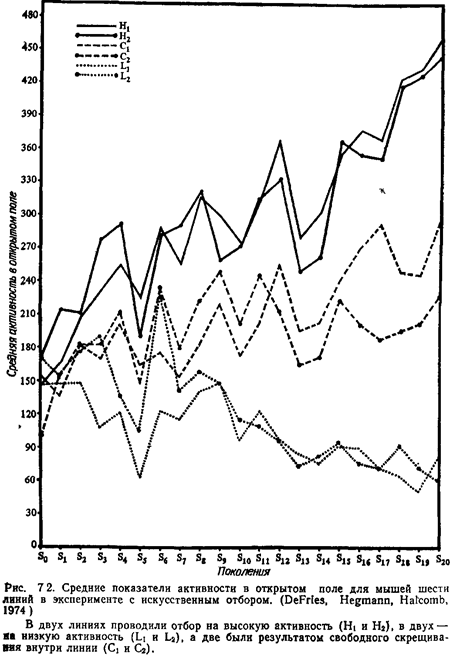
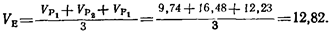
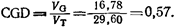

 Рис 85 Чувствительный период для импринтинга. (Hess, 1959)
Рис 85 Чувствительный период для импринтинга. (Hess, 1959)