|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Глава 7 ГЕНЕТИКА ПОВЕДЕНИЯ
Ученые давно интересовались и теориями, и фактическими данными относительно роли наследственных факторов в поведении. Подобный интерес можно проследить со времен древних греков, затем у Леонардо, а далее у Монтеня, Дарвина, Гальтона, Иеркса и Трайона (McClearn, DeFries, 1973). Однако как отдельная дисциплина генетика поведения совсем еще молода. Заслуга выделения ее как четко определенной области исследования принадлежит Джону Фуллеру и У. Р. Томсону, которые в 1960 г. опубликовали свою монографию «Генетика поведения» («Behavior genetics»). В последние 20 лет изучение влияния генетических факторов на поведение быстро продвигалось вперед. Поскольку разные исследователи интересуются разными проблемами, работы в этой области удобно разбить на шесть групп, соответствующих следующим вопросам (по Thiessen, 1972, видоизменено): 1. Влияют ли различия в генотипе на исследуемое поведение? 2. Если влияют, то сколько генов к этому причастно? 3. Какая доля наблюдаемой изменчивости поведения определяется генетическими и какая – средовыми факторами? 4. Какие онтогенетические и физиологические процессы опосредуют влияние генов на поведение? 5. Что может дать генетический анализ для выяснения возможной адаптивной роли данной формы поведения? 6. Как взаимодействуют гены и внешняя среда в детерминации данного поведения? В настоящей главе будут рассмотрены эти шесть фундаментальных вопросов, методы, которые используются при их изучении, и некоторые уже найденные ответы. При обсуждении данных по генетике поведения нам придется использовать ряд основных генетических понятий. Часть из них уже была разъяснена в главе 3; возможно, читателю полезно будет еще раз вернуться к этому месту, прежде чем идти дальше.
ВЛИЯЕТ ЛИ НА ПОВЕДЕНИЕ ГЕНОТИП? По мнению сторонника одних лишь средовых влияний, гены фактически не играют никакой роли в контроле поведения. Сегодня такую позицию почти все считают неверной. Существует множество убедительных данных о влиянии генотипа на поведение, хотя все они в известной степени непрямые. Фактически все исследования, о которых будет идти речь в этой главе, приводят к одному и тому же выводу: изменения в генотипе глубоко влияют на внешне наблюдаемое поведение. Эти данные могут убедить даже самого непримиримого скептика. Вся эта глава посвящена вопросу о генетических влияниях на поведение; мы начнем с того, что рассмотрим три пути подхода к его решению: изучение межлинейных различий, опыты с отбором и межвидовую гибридизацию. Межлинейные различия Основной принцип использования межлинейных различий для решения интересующего нас вопроса очень прост. На развитие поведения влияют два главных фактора – гены и среда. Стратегия исследования состоит в том, чтобы найти организмы, различающиеся по генотипу, и, поддерживая постоянство внешних условий, определить, влияют ли на поведение генетические факторы. Главная трудность генетического исследования связана с тем, что гены нельзя наблюдать непосредственно. Для того чтобы делать выводы относительно генотипа, приходится применять косвенные методы. Как же в таком случае мы можем найти организмы с различным генотипом? Многие исследователи используют для этого инбредные линии. В инбредной линии все особи фактически гомогенны, т. е. генетически идентичны, и гомозиготны по всем локусам. Инбредные линии домовой мыши (Mus musculus) – наиболее -популярного объекта в исследованиях по генетике поведения млекопитающих – выводят путем скрещивания братьев с сестрами на протяжении по меньшей мере 20 поколений. Из генетически гетерогенной популяции берут брата и сестру и получают от них потомство. Брата и сестру из этого потомства снова скрещивают между собой для получения следующего поколения, и т. д. С каждым поколением данная линия безвозвратно теряет некоторые аллели, и примерно к 20-му поколению она приближается к полностью изогенному состоянию. Отметим, что в этом случае животных для скрещивания подбирают на основе их генетического родства – в отличие от селекции на определенные признаки (анатомические, физиологические или поведенческие), о которой речь пойдет позже. В значительной мере благодаря большой программе, осуществленной в Джексоновской лаборатории (штат Мэн, США), в настоящее время исследователи располагают большим числом инбредных линий животных. Каждая из них получила свое обозначение в соответствии с международной системой (например, C57BL/6J). Если животные разных инбредных линий, выращенные в идентичных условиях среды, обнаруживают существенные различия, то весьма вероятно, что эти различия имеют генетическую основу. Подобные результаты были получены для целого ряда особенностей поведения, включая активность в «открытом поле» (Thompson, 1953), агрессивное поведение (Southwick, Clark, 1968), половое поведение (McGill, 1962), постройку гнезда (Lynch, Heg-mann, 1972) и научение (Wahlsten, 1972). Фактически даже труднее найти линии, которые не отличались бы друг от друга, чем линии, между которыми есть различия. Спрот и Стаатс (Sprott, Staats, 1975) составили полную библиографию (1222 названия) работ по поведению с использованием генетически охарактеризованных мышей. Перекрестное воспитание. Хотя межлинейные различия указывают на генетический эффект, не исключено, что эти различия обусловлены какими-то тонкими средовыми факторами. Один из способов контроля таких факторов – эксперимент с перекрестным воспитанием. Пометы новорожденных мышат, принадлежащие к линиям с разным поведением во взрослом состоянии, меняются местами, так что матери одной линии воспитывают детенышей другой, и наоборот. Контрольные животные воспитываются либо собственными матерями, либо другими самками той же линии. Один эксперимент такого рода был проведен Саутвиком (Southwick, 1968). Линии A/J и CFW сильно различались по агрессивному поведению. Полученные результаты представлены на рис. 7.1. Воспитание мышей A/J матерями из линии CFW существенно повышало показатель их агрессивности. Однако это изменение невелико по сравнению с уровнем межлинейных различий.
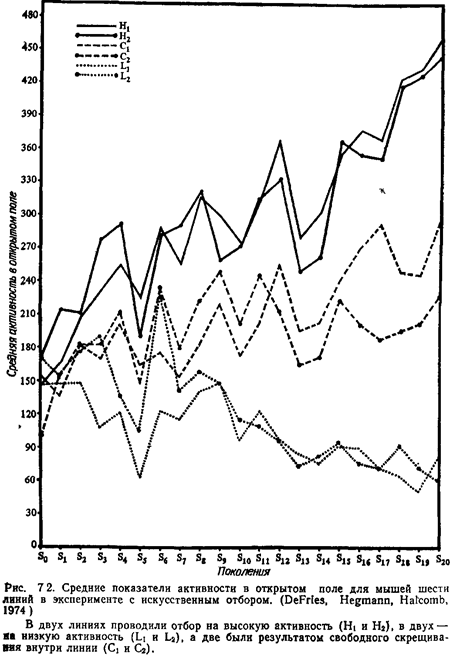
Воспитание мышей CFW матерями A/J не влияло на агрессивность потомства. Таким образом, различия между этими линиями в агрессивном поведении, по-видимому, обусловлены в основном наследственностью. Трансплантация яичников. Эксперимент с перекрестным воспитанием устраняет не поддающиеся контролю постнатальные влияния со стороны матери, однако пренатальные средовые условия при этом не изменяются. Метод трансплантации яичников позволяет отделить генетические эффекты от пренатальных материнских влияний. У животных инбредных линий происходит отторжение тканей, пересаженных им от животных других линий. Если, однако, скрестить две инбредные линии и получить гибридов первого поколения (Fi), то у этих гибридов трансплантаты яичников от обеих родительских линий не отторгаются. Скрещивая самок Fi, имеющих пересаженные «инбредные» яичники, с самцами той же инбредной линии, можно затем сравнить поведение мышей двух разных линий, развивавшихся либо у инбредных матерей, либо у матерей Fi. Используя этот метод, Де-Фриз и др. (DeFries et al., 1967) нашли, что в детерминации различий поведения в открытом поле главную роль играют генетические факторы, в то время как вес тела больше зависит от материнского влияния. Эксперименты с отбором Работы по искусственному отбору отличаются от попыток создать инбредные линии тем, что животных для скрещивания отбирают не на основе генотипа, а по фенотипу. Фенотипический признак – это любой доступный для измерения морфологический, физиологический или поведенческий признак организма. Вес тела, интенсивность обмена, активность в открытом поле – все это фенотипические признаки. Генотипом называют весь генетический материал организма или его часть. Для того чтобы начать эксперимент с искусственным отбором, нужно определить у группы самцов и самок из генетически гетерогенной популяции какой-либо признак. Самцов с высокими величинами данного признака скрещивают с такими же самками, а самцов с низкими величинами – с самками, у которых этот признак тоже слабо выражен. Потомство от скрещиваний тестируют на степень проявления того же признака, и для следующего скрещивания пары снова подбирают по такому же принципу. Таким образом в последовательных поколениях получаются две линии – одна в результате отбора на высокие показатели данного признака, а другая – на низкие. Для контроля изменений в условиях среды ведут еще третью линию – путем случайного скрещивания. Если в определении изучаемого признака имеется генетический компонент, показатели «высокой» и «низкой» линии в последовательных поколениях расходятся. Показатели линии со случайным скрещиванием не должны обнаруживать систематических отклонений. Де-Фриз, Хегман и Халком (DeFries, Hegmann, Halcomb, 1974) использовали метод отбора для анализа поведения мышей в открытом поле. Взяв в качестве исходного материала относительно гетерогенную группу, они в течение 20 поколений вывели две линии с высоким значением признака, две линии – с низким и две контрольные линии. Результаты представлены на рис. 7.2. Можно видеть, что за эти 20 поколений показатели признака для различных линий разошлись – они увеличились у животных с высоким значением признака и снизились в линии с низким значением, а у контрольных линий изменились очень мало. Эти данные иллюстрируют важную роль генов в контроле активности животных в открытом поле. Сходные результаты были получены в селекционных экспериментах с широким спектром видов и фенотипиче-ских признаков, включая фототаксис (тенденцию приближаться к источнику света) у плодовой мушки (Hirsch, Boudreau, 1958), научение крыс в лабиринте (Тгуоп, 1940), реакции, связанные с импринтингом, у цыплят (Graves, Siegel, 1969) и агрессивность у мышей (Lagerspetz, 1969).
Межвидовые гибриды В большей части исследовании по генетике поведения используют особей одного вида, имеющих разные генотипы. Однако в лабораторных условиях можно скрещивать особей разных, но близких друг к другу видов. В поведении потомков от такого скрещивания часто обнаруживаются явные признаки генетических влияний. Превосходный пример такого подхода – работа Дилгера (Dilger, 1962), который изучал поведение, связанное с постройкой гнезда, у разных видов неразлучников. Особи одного вида, Aga-pornis personata, отрывают кусочки материала для гнезда и переносят их к гнезду в клюве. Представители другого вида, А. го-seicollis, не держат их в клюве, а засовывают под боковые перья. Исследованные Дилгером гибриды f[ обнаружили смешанный тип поведения, при котором птицы пытались засунуть материал для гнезда в перья, затем вынимали его, брали их в клюв, а потом все начиналось сначала. Таким образом, их поведение представляло собой малоэффективную смесь из двух типов целесообразного поведения родительских форм. Сходные промежуточные формы обнаруживались в демонстрационном поведении вьюрков (Hinde, 1956) и в характере звуковых сигналов сверчков (Bentley, Hoy, 1974).
|
Последнее изменение этой страницы: 2019-04-09; Просмотров: 355; Нарушение авторского права страницы