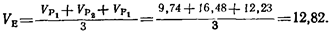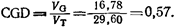|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Плейотропия и признаки с непрерывным распределением
Все признаки, которые мы до сих пор рассматривали при анализе моногенного наследования, характеризовались дискретным распределением. В этом случае каждую особь по ее фенотипу можно отнести к одной из нескольких обособленных категорий, а промежуточные фенотипы не встречаются. Примером дискретного признака служит пол. Особь может быть либо самцом, либо самкой – промежуточные формы очень редки. Устойчивость к аудиогенным судорогам, писк мышей, ощущение вкуса фенилтио-карбамида, гигиеническое поведение – всем этим признакам свойственно дискретное распределение. Поскольку мы можем относить особей к четко разграниченным группам, проводить генетический анализ сравнительно просто. К сожалению, для большинства признаков, касающихся поведения животных, характерно непрерывное распределение. По степени проявления такого признака особь может находиться в любой точке некоторого континуума. Размеры и вес тела, число квадратов, посещаемых в тесте с открытым полем, время возвращения детеныша в гнездо – все это признаки с непрерывным распределением. Можно ли в признаке с непрерывным распределением обнаружить влияние отдельных генов? Ответ будет утвердительным, однако при этом возникает ряд трудностей. Одна из них – проблема идентификации особей, у которых есть интересующий нас ген. Так как мы не можем видеть гены непосредственно, мы должны полагаться на косвенную оценку генотипов. Наиболее распространенный метод основан на явлении плейотропии. Так называют одновременное влияние гена на несколько различных признаков. Наличие или отсутствие гена можно установить по одному из его плейотропных эффектов. Затем можно перейти к поиску сопутствующего влияния этого гена на поведение. Например, у мышей альбинизм контролируется одним рецессивным геном. Было показано, что ген альбинизма влияет на некоторые поведенческие признаки; в частности, у мышей, имеющих этот ген, понижена активность в открытом поле (DeFries, 1969). При работе с дрозофилой можно применять и более сложные методы выявления эффектов единичных генов (см., например, Kessler, Kraemer, 1975). Среди исследователей нет согласия относительно того, имеет ли смысл искать эффекты отдельных генов, изучая признаки с непрерывным распределением (см., например, Wilcock, 1969; Thie-ssen, 1971). По-видимому, большая часть признаков, представляющих интерес для исследователей поведения животных, имеет непрерывное распределение и детерминирована действием многих генов, каждый из которых сравнительно мало влияет на фенотип.
КАКОВ ОТНОСИТЕЛЬНЫЙ ВКЛАД ГЕНЕТИЧЕСКИХ И СРЕДОВЫХ ФАКТОРОВ В НАБЛЮДАЕМУЮ ИЗМЕНЧИВОСТЬ? Исследуя признаки с непрерывным распределением, зависящие от многих генов, необходимо применять методы биометрической, или количественной, генетики (см. Falconer, 1960). Эти методы позволяют оценивать ряд характеристик или параметров генетической структуры, которые влияют на интересующую нас особенность поведения. Однако расчеты основываются- здесь на ряде допущений -относительно свойств генов и шкалы измерения зависимой переменной. Поскольку при работе с данными о поведении эти допущения не всегда легко оправдать, при рассмотрении некоторых более сложных моделей вполне уместен известный скептицизм. Наследуемость В генетике поведения большое значение придается вычислению наследуемости. Эта величина говорит о том, какую долю общей изменчивости данного признака (в популяции) можно отнести за счет различий в генотипе. Нам нужно будет сначала рассмотреть метод оценки изменчивости – изменение вариансы. Варианса служит показателем дисперсии (или изменчивости) выборки. Предположим, что у нас есть две группы животных по три особи в каждой; животные одной группы посетили соответственно 49, 50 и 51 квадрат открытого поля, а животные другой группы – 30, 50 и 70 квадратов. Средние для обеих групп между собой равны, однако группы различаются по вариабельности отдельных значений относительно средней: во второй группе изменчивость больше. Варианса позволяет оценивать эту изменчивость количественно. Варианса – это средняя величина квадратов отклонений от среднего значения признака. Для вычисления вариансы находят разности между каждой величиной и средней, возводят их в квадрат и находят среднюю от этих квадратов. Вариансы двух приведенных выборок равны соответственно 0, 7 и 266, 7. Следует вспомнить, что всех особей одной инбредной линии можно считать генетически идентичными. Таким образом, в инбредной линии отсутствует генетическая варианса, поэтому как vq, так и CGD должны быть равны нулю Это не означает, что на данный признак генотип не влияет, а значит лишь, что в данной группе животных нет той доли изменчивости, которая определялась бы различиями в генотипе Если бы мы могли выращивать группу генетически гетерогенных особей в совершенно одинаковых условиях, CGD приближалось бы к 1. Это не значит, что средовые факторы для развития данного признака несущественны, а означает лишь, что в данной группе и в данных условиях различия между особями не определяются средой. Если мы обозначим генетическую аддитивную вариансу как va, то истинную наследуемость, или наследуемость «в узком смысле» (/i2), можно будет представить следующей формулой: Различие между va и vg состоит в том, что аддитивная варианса va. – это только часть всей генетической вариансы vg. Часть общей генетической вариансы связана с действием доминантных генов, подавляющих проявление своих рецессивных аллелей, а часть связана с эпистазом, т. е. взаимодействием между генами разных локусов Поскольку эта часть генетической вариансы следующему поколению непосредственно не передается, при статистической обработке ее исключают, чтобы получить величины аддитивной генетической вариансы и истинной наследуемости. Величина h2 всегда должна быть равна или меньше величины CGD, поскольку Уд – только часть общей генетической вариансы Ve-«Наследуемость признака – один из самых важных популяцион-ных параметров, которые можно оценить методами количественной генетики» (DeFries, Hegmann, 1970, р. 27). Вычисление наследуемости Наследуемость или CGD можно вычислять многими способами. Для людей можно использовать результаты близнецового анализа, корреляции между родителями и детьми и между сибсами. В исследованиях, проводимых на животных, обычно используют метод отбора, менделевские скрещивания или диаллельное скрещивание (см. ниже), чтобы находить величины h2 или CGD. У Фуллера (Fuller, 1960) мы находим пример вычисления CGD на основе данных менделевского скрещивания при изучении исследовательской ^активности мышей, помещенных в лабиринт Полученные данные представлены в табл. 7 2. Здесь приводятся средние и вариансы величин исследовательской активности для двух инбредных линий, BR и А, и для их гибридов Fi и F2 Для оценки CGD нам нужно знать величины оценок vq и Ут. Поколение Fz содержит практически все генотипы и поэтому поможет нам получить оценку V^. Поскольку все особи в каждой из инбредных линий и в гибридном поколении Fi генетически идентичны друг другу (все животные Fi гетерозиготны по всем локусам, по которым различаются инбредные линии), генетической вариансы внутри каждой из этих групп не будет.
Таблица 7.2 Исследовательская активности у мышей линий C57BR и A/Jax и их гибридов (Fuller, 1960)
Каждая из получаемых величин вариансы будет оценкой ve- Наилучшей оценкой ve будет средняя для трех варианс:
Генетическую вариансу Vo можно найти, вычтя ve из V-s (vg^v-s – ve; 29, 60 – 12, 82=16, 78). Тогда
Таким образом, мы установили, что несколько больше половины вариансы исследовательской активности у этих групп связано с генетическими факторами. Сходные величины были получены при исследовании мышей линий C57BL и BALB/c (DeFries, Hegmann, 1970). Хотя этот метод дает хорошую аппроксимацию величины CGD, его часто не удается применить в исследованиях по генетике поведения. Дело в том, что варианса Fz нередко бывает не выше вариансы Fi или родительских линий – так называемый «эффект Трайона» (Hirsch, 1967; DeFries, Hegmann, 1970). Это, видимо, связано с тем, что признаки поведения обычно находятся под влиянием большого числа генов. Наследуемость или CGD определяли, используя множество линий, методов и поведенческих признаков. Фуллер и Уаймер (Fuller, Wimer, 1973) подвели итог некоторым результатам; оказалось, что эти величины варьируют в пределах от 0, 22 для агрессивности у кур (оценка с использованием метода отбора) до 0, 89 для половой активности морских свинок (по данным скрещивания инбредных линий). |
Последнее изменение этой страницы: 2019-04-09; Просмотров: 323; Нарушение авторского права страницы