
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Импринтинг (запечатление)⇐ ПредыдущаяСтр 11 из 11
Поведенческие акты птиц в природе адресуются другим организмам не случайным образом – обычно они направлены на биологически адекватные мишени. В своих вылазках из гнезда в ранний период жизни птенцы выводковых птиц следуют за матерью. По достижении половой зрелости большинство птиц выбирает брачного партнера только среди особей своего вида. Вопрос о природе процессов, обеспечивающих такую избирательность поведения, привлекал большое внимание исследователей. История вопроса. Ученые нашли много ранних упоминаний о явлениях, подобных импринтингу (например, Kevan, 1976). Из ранних работ наиболее содержательным было исследование Сполдин-ra (Spalding, 1873), который среди других явлений заметил, что, как только цыплята приобретают способность ходить, они начинают следовать за любым движущимся объектом. Хейнрот расширил наблюдения подобных явлений, исследуя множество разных видов птиц. Интерес к импринтингу больше всего стимулировали работы Конрада Лоренца (Lorenz, 1937a). Этот автор описал многочисленные случаи, когда птицы не спаривались с особями своего вида из-за того, что они были лишены контакта с ними в ранний период жизни. Например, Лоренц выращивал гусей в изоляции от сородичей в течение первой педели жизни и отметил, что такие птицы предпочитают следовать за людьми, а не за другими гусями. Птицы ждали перед парадной дверью дома Лоренца и пытались следовать за людьми, как только те выходили из дома. Лоренц назвал это явление Pragung, что по-немецки означает «впечатывание», а на английский было переведено как imprinting (запечатление). Лоренц высказал предположение, что импринтинг отличается от истинного ассоциативного научения четырьмя особенностями: 1. Импринтинг приурочен к очень ограниченному периоду жизни птицы – «критическому периоду». 2. Однажды совершившись, процесс запечатления далее необратим. Если у птенца произошел импринтинг по отношению к птице другого вида, то позднейший контакт с особями своего вида уже не сможет устранить этот эффект раннего опыта. 3. Импринтинг – уникальный феномен в том отношении, что объект, на который будет направлено поведение, может быть определен задолго до первого осуществления самого поведения. Было, например, показано, что ранний импринтинг, происходящий задолго до наступления половой зрелости, влияет на выбор полового партнера. 4. Лоренц рассматривал импринтинг как форму «супериндивидуального условного рефлекса» в том смысле, что поведение при этом будет направлено не на определенную особь, которую видел птенец, а на целый класс организмов, членом которого была особь, послужившая стимул-объектом. Здесь следует отметить, что в дальнейшем многие исследователи подвергли сомнению, действительно ли все эти отличия столь существенны Начиная с 1937 года импринтинг зарекомендовал себя как увлекательная проблема и для психологов, и для зоологов, которые в попытках понять это явление потратили немало часов и исписали немало страниц. Поскольку в работах по половому импринтингу и но запечатлснию привязанности использовались несколько различные методы и получены разные результаты, мы рассмотрим эти работы раздельно Запечатление привязанности. Говоря об этом явлении, мы рассмотрим проблемы, связанные с механизмом реакции следования у выводковых птиц. Говоря проще, нас интересует, как молодая птица узнает, за кем она должна следовать.
Рис 8 4 Аппарат для лабораторных опытов по изучению импринтинга (Hess, 1959) Птенец попадает на круговую дорожку и может видеть модель, движущуюся с постоянной скоростью. Методы. Большая часть исследований по запечатлению привязанности была выполнена в лабораториях, например в лаборатории Экхарда Хесса (Hess, 1959). Основной аппарат Хесса изображен на рис. 8.4. Птенец вылупляется из яйца в темноте, и его содержат изолированно в темном ящике внутри инкубатора вплоть до самого момента опыта. Через автоматическую дверцу птица попадает в аппарат на круговую дорожку диаметром около 1, 5 м. В большинстве экспериментов по дорожке с определенной скоростью перемещали чучело селезня (рис. 8.4), зеленую коробочку или красный мяч. В некоторые объекты был вмонтирован динамик, так что они могли издавать звуки. Птенец оставался в аппарате определенное время – на период запечатления. Если соблюдены все условия, он начинает следовать за движущимся объектом. Из аппарата птенца извлекают через дверку в полу. Позже его снова помещают в аппарат на время тестирования. При этом птенцу обычно предъявляют на выбор два объекта: тот, который ему ранее показывали, и какой-нибудь новый. Обычно птенец оказывает предпочтение объекту, который уже экспонировался. С помощью описанного аппарата можно проделывать много различных манипуляций Рассмотрим результаты некоторых экспериментов.
Стимул-объекты. Мозг птенца не tabula rasa (чистая доска) в том смысле, который придавал этому выражению Джон Локк. Хотя в экспериментах с имприитингом можно успешно применять Представлен процент птиц, обнаруживающих наилучшие показатели запечатления, как функция возраста.при первом предъявлении стимул объекта. самые различные стимул-объекты, некоторые из них более эффективны, нежели другие (Bateson, 1971; Graves, 1973). Стимулы, более заметные для человеческого глаза, оказываются более эффективными и для импринтинга. Например, Клопфер (Klopfer, 1967) нашел, что запечатление бывает значительно сильнее, когда используется модель, разрисованная в полоску, по сравнению с моделью, выкрашенной белой краской. Оказалось, что птицы, следовавшие в период импринтинга за белой моделью, в период тестирования предпочитали ей полосатую. Таким образом, хотя вообще птицы предпочитают следовать за объектами, на которые у них произошел импринтинг, мощные зрительные раздражители могут перекрывать эффект раннего опыта. Чувствительный период. В представлениях Лоренца об имприн-тинге важнейшую роль играл «критический период». В целом экспериментальные данные подтверждали, что наиболее эффективное запечатление происходит во время че'''ко ограниченной фазы развития организма. На рис. 8 5 представлены результаты, полученные Хессом. У утят готовность к импринтингу достигает максимума в период от 13 до 16 часов жизни. Воздействие стимул-объектов в более ранние и более поздние сроки неэффективно. Под «критическим периодом» обычно понимают тот период онтогенеза, когда определенная система наиболее подвержена модификации. Вероятно, в такие периоды происходит наиболее быстрое созревание механизмов, необходимых для данной реакции. Концепция критического периода вызвала много споров. Фактически все исследователи согласны в том, что запечатление. например, не происходит одинаково хорошо на всех стадиях жизни. Однако некоторые авторы утверждают, что изменения в чувствительности происходят несколько более плавно, чем можно было бы ожидать, исходя из концепции критического периода или из данных, представленных на рис. 8 4. Кроме того, оказалось, что для ряда поведенческих систем можно изменять временные характеристики «критического периода» Например, на период чувствительности к импринтингу могут влиять условия выращивания (Bateson, 1971). В связи с этим многие исследователи стали называть этот период не «критическим», а «чувствительным», чтобы не создавать представления о внезапности и жесткой фиксирован-ности происходящих изменений. Этот менее спорный термин мы и будем использовать. Однако это не должно умалять значение концепции об ограниченном периоде максимальной готовности к импринтингу. Запечатленные объекты как подкрепляющие стимулы. Показано, что объекты, на которые произошел импринтинг, служат для молодых птиц эффективными подкрепляющими стимулами. Если предъявлять такой объект после какой-либо произвольно выбранной реакции, птицы могут обучиться этой реакции Например, Хофман и др. (Hoffman et al, 1966) нашли, что цыплята клюют рычаг, если эта реакция ведет к появлению перед ними запечатленного объекта – пластиковой молочной бутылки. Процессы, лежащие в основе импринтинга. Вопрос о природе механизмов, лежащих в основе импринтинга, привлекал большое внимание и вызывал много споров. Недостаток места не позволяет нам рассмотреть этот вопрос подробно. Ражецкий (Rajecki, 1973) приводит шесть разных точек зрения относительно механизмов импринтинга: это интерпретации с точки зрения контекстуальной, этологической, условно-рефлекторной, эпигенетической и информационной концепций, а также концепции нейронных цепей Обратимость. Так же как и чувствительный период, свойство необратимости играло важнейшую роль в концепции импринтинга, предложенной Лоренцем. Поскольку содержать большое число животных в течение долгого времени довольно дорого, обратимость запечатления привязанности была предметом лишь немногих длительных исследований. Видимо, она варьирует в зависимости от природы раздражителя и условий импринтинга. В большинстве случаев запечатление привязанности, изучаемое в лабораторных условиях, оказывается легко обратимым Например, Хесс (Hess, 1972) показывал маленьким утятам людей в течение 20 часов непрерывно. Утята следовали за людьми всюду Однако они стали следовать за живой уткой-кряквой, после того как ее показывали им в течение полутора часов в более позднем возрасте. «Чисто лабораторный импринтинг обратим», – заключает автор (Hess, 1972, р. 25). И наоборот, Хесс не смог изменить импринтинг, который произошел у утят на их собственную мать-утку, которую они видели в течение 16 часов сразу после вылуп-ления. Межвидовые различия. Следует отметить, что разные виды в сходных ситуациях реагируют по-разному. Хотя очевидно, что у домашних цыплят и у утят кряквы закономерности импринтинга могут быть различными, игнорирование таких различий было причиной некоторой путаницы в литературе по импринтингу. Роль слуховых стимулов. Уже давно известно, что комбинированное воздействие зрительных и слуховых раздражителей при импринтинге более эффективно, чем действие одних только зрительных стимулов. Готлиб (Gottheb, 1973) сообщил о серии экспериментов, направленных на выяснение относительной важности тех и других раздражителей. Он исследовал птиц, которые вылуплялись и далее выращивались в условиях, когда только слышали, но не видели своих сибсов и никогда не видели взрослых особей и не слышали издаваемых ими звуков. Маленькие утята трех видов с готовностью следовали за моделью, похожей на утку-мать соответствующего вида, издававшей такие же звуки, как эта утка. Однако подмена зрительных компонентов такого стимул-объекта зрительным образом, характерным для другого вида, на реакцию следования почти не влияла. Напротив, изменение звукового сигнала существенно ослабляло реакцию следования. Птицы, никогда не слышавшие материнского зова, избирательно реагировали на крики особей своего вида Например, птенцы пекинской утки вначале следовали за одной из двух идентичных моделей матери, а затем, когда звуковые сигналы, издаваемые «матерями», менялись местами, утята либо изменяли предпочтение, либо просто переставали следовать за первой моделью. Эти наблюдения позволяют предполагать, что в ранний период жизни утят слуховая избирательность может быть первичной детерминантой реакции следования. Маленькие утята «попадают в ситуацию импринтинга уже „настроенными" на определенные аудиометрические характеристики материнского призывного сигнала своего вида» (Gottlieb, 1973, р. 371). Хесс показал, что между матерью-уткой и эмбрионом непосредственно перед вылуплением протекают настоящие диалоги. Хесс предполагает также, что таким путем молодые птенцы могут усвоить определенные звуковые характеристики сигналов именно их матери и таким образом окажутся способными отличать ее сигнал от сигналов птиц других видов Половой импринтинг. Говоря о «половом импринтинге», мы имеем в виду процесс, в результате которою данное животное начинает адресовать свое половое поведение одним особям, а не другим Проще говоря, птица должна определить, с кем ей надлежит спариваться Часто предполагают, что половой импринтинг и запечатление привязанности происходят в результате одного и того же процесса в те же самые сроки. Однако дело, видимо, не всегда обстоит так. Иногда эти два процесса могут быть разделены. Методы. Методы исследования полового импринтинга отличаются от тех, которые используют в случае запечатления привязанности. ^Обычно длительность экспозиции объекта в исследованиях по половому импринтингу значительно больше. Исследуя внутривидовой половой импринтинг, птицу выращивают с особями того же вида, но отличающимися по окраске (например, белыми). Такие эксперименты проводились с разными видами птиц – с утками, голубями, вьюрками. При изучении межвидового импринтинга птиц выращивают с особями других видов. Такие работы были проделаны с различными представителями утиных, куриных и воробьиных. Следует отметить, что здесь можно использовать птиц как выводковых, так и птенцовых видов. Особую категорию составляют эксперименты, в которых объектом запечатления служит человек. Хотя некоторые из подобных данных можно считать всего лишь анекдотическими, в настоящее время половой импринтинг на человека у выращенных в неволе птиц описан у 25 видов (Immelmann, 1972). При изучении полового импринтинга пробу на предпочтение полового партнера проводят в зрелом возрасте. Некоторые исследователи для ускорения полового созревания вводили птицам гормоны, однако такие процедуры могут исказить нормальный ход процессов развития. Стимул-объекты. Успешность полового импринтинга сильно варьирует в зависимости от использованного объекта. В некоторых работах, например, импринтинг на человека оказывается несколько более слабым, чем на особей другого вида, более похожих на птиц данного вида и поведением, и внешностью. Исследования по половому импринтингу на неодушевленные предметы, кажется, почти не проводились. Межвидовые различия. У разных видов относительная роль врожденных перцептивных образов и научения в феномене импринтинга весьма различна (Lorenz, 1937a, р. 268). Способность к запечатлению может варьировать внутри вида в зависимости от пола. Чувствительные периоды. Чувствительные периоды для полового импринтинга, по-видимому, менее жестко ограничены во времени и часто приходятся на несколько более поздний период жизни, чем периоды для запечатления привязанности. В большинстве исследований, в которых был обнаружен сильно выраженный половой импринтинг, экспозиция была сравнительно долгой. Половой импринтинг может происходить в совершенно иной период, нежели запечатление привязанности. По данным Иммельмана (Immelman, 1972), у серого гуся чувствительный период начинается около 50-дневного возраста и кончается примерно к 140-му дню, а у крякв, видимо, начинается между 5 и 19 днями жизни и длится несколько недель. Клингхаммер (Klinghammer, 1967) отмечает, что у галок, зебровых амадин и дубоносов половой импринтинг происходит относительно рано в гнездовой период, а у снегирей, ворон и воронов окончательный выбор полового партнера определяется перед достижением половой зрелости. У горлиц и других голубей на выбор полового партнера можно влиять еще долго после того, как молодые птицы становятся независимыми от родителей. Обратимость. Хорошо известно, что птицы, у которых произошел импринтинг на особей другого вида, могут спариваться также и с партнерами своего вида, если тестирование производится каждый раз только с одним из этих видов. Если же птице предоставить выбор, она часто проявляет предпочтение, которое, видимо, вполне устойчиво. Например, Шейн (Schein, 1963) исследовал трех индюков, которые в течение первых 32 дней жизни росли в изоляции от своего вида, а в возрасте 5 лет тестировались на предпочтение полового партнера. Птицы «ухаживали» за человеком в присутствии людей и особями своего вида, когда находились только с ними. В ситуации выбора они ухаживали только за людьми. Хотя в зависимости от длительности контакта, природы стимул-объекта и изучаемого вида результаты бывают разными, половой импринтинг все же оказывается достаточно устойчивым. Импринтинг у птиц. Обобщение. Литературу по импринтингу следовало бы рассмотреть подробнее. Это процесс очень сложный. Некоторые авторы склонны проводить аналогию между имприн-тингом у птиц и явлениями научения в раннем возрасте у других видов, в особенности у детей. При проведении таких аналогий запечатление у птиц обычно рассматривается как процесс, протекающий в пределах жестких сроков критического периода в раннем возрасте, совершающийся практически мгновенно наподобие «впечатывания» и совершенно необратимый. На самом же деле в зависимости от вида птиц, от стимул-объектов, типа изучаемого импринтинга и многих других факторов при запечатлении обнаруживаются совершенно различные функциональные особенности. При этом можно продемонстрировать полную гамму явлений, начиная от отсутствия предпочтения запечатленного объекта до аномального полового импринтинга, эффект которого сохраняется всю жизнь. Как в случае почти всякого " хорошо изученного поведенческого феномена, детерминанты оказываются сложными. Поспешное распространение выводов на другие виды и другие ситуации было бы здесь неоправданным. Импринтинг у млекопитающих. Явления, очень сходные с запе-чатлением привязанности у птиц, продемонстрированы у некоторых зрелорождающихся млекопитающих. Например, Слакин (Sluckin, 1968) смог показать, что у морских свинок обнаруживается дифференциальное предпочтение кубика, окрашенного в белый и черный цвета, по сравнению с теннисным мячом в зависимости от того, какой из этих предметов предъявлялся им в раннем возрасте. Клопфер (Klopfer, 1971) исследовал другой феномен – «материнское запечатление». Козы-матери избирательно реагируют на своих собственных детенышей, предпочитая их детенышам других самок. Все 15 коз, у которых детенышей отобрали сразу после родов, отказывались принять козлят, когда их возвращали спустя 1 – 3 часа. Напротив, 14 из 15 матерей, которым в сходном опыте был предоставлен пятиминутный период контакта с детенышами, козлят приняли. По-видимому, материнское запечатление – процесс очень быстрый. Влияние раннего опыта у млекопитающих В последнюю четверть века мы были свидетелями настоящего взрыва исследований по влиянию раннего опыта на последующее поведение млекопитающих. Целая армия психологов, работающих в области экспериментальной и сравнительной психологии, а также психологии развития, произвела на свет буквально сотни оригинальных и обзорных статей на эту тему. Стимулом для этого служили интересные данные об импринтинге у птиц, некоторые приложения теории Фрейда, а также очевидная важность сведений о влиянии условий воспитания в раннем возрасте на поведение взрослого человека. В этих работах использовались самые различные виды животных и типы экспериментальных воздействий. Поскольку на нескольких страницах невозможно дать полный обзор таких исследований, мы попытаемся лишь дать о них общее представление. Мы рассмотрим данные о влиянии некоторых воздействий на животное в пренатальном периоде, обогащения среды в раннем возрасте и депривации в этот же период. Пренатальные воздействия. Чтобы исследовать влияние пре-натальных факторов, беременную самку обычно подвергают действию того или иного стресса. Во многих случаях это приводило к стойким изменениям в поведении потомства. Наиболее распространенная гипотеза относительно механизмов такого эффекта состоит в предположении, что агенты, вызывающие стресс, изменяют гормональную среду организма самки, а это влияет на плод таким образом, что последствия проявляются на более позднем этапе жизни. Томпсон (Thompson, 1957) воздействовал на группу самок сочетанием звонка и электрического удара. Они научились избегать удара, перебегая на безопасную половину челночной камеры при звуке звонка. Во время беременности самок три раза в день подвергали действию звонка, но не давали ни тока, ни возможности осуществить реакцию избегания. Для исключения возможных постнатальных влияний потомство воспитывалось другими самками. Оказалось, что пренатальный стресс матери снижает уровень активности и удлиняет латентный период ее начала в тесте с «открытым полем»; удлиняется и латентный период выхода из обычной клетки для побежки за пищей после 24-часового голодания. Эти изменения были истолкованы как результат повышенной «эмоциональности» потомства тех матерей, которые были подвергнуты стрессу. Как и во многих других случаях, гены и внешняя среда здесь взаимодействуют, формируя определенный результат эксперимента. Де Фриз, Вейр и Хегман (DeFries, Weir, Hegmann, 1967) показали, что влияние физического или химического стресса, которому подвергали беременных мышей-самок, на поведение их потомства в тесте открытого поля зависит от линии отца, линии матери и типа стрессорного воздействия. Уорд (Ward, 1972) нашел, что стресс матерей в пренатальный период снижает у крыс-самцов копулятивную активность и способствует проявлению у них реакции лордоза, характерной для самок. Раннее обогащение среды. Огромное большинство работ по влиянию раннего опыта у млекопитающих касалось последствий обогащения среды в раннем возрасте. В лабораториях грызунов чаще всего содержат в относительно небольших стерильных металлических клетках. Под «обогащением» среды мы понимаем введение каких-либо факторов, дополняющих или усложняющих -обычную среду в условиях лаборатории. Литература по этому вопросу очень сложна, так как почти всем опытам, в которых обнаруживается тот или иной эффект, можно противопоставить другие опыты, в которых сходное воздействие этого эффекта не дает или даже вызывает противоположное изменение. Это обусловлено несколькими причинами. Основная трудность связана с большим числом переменных, которые могут влиять на поведение в более позднем возрасте, и с тем, что в разных экспериментах было использовано значительное число различных параметров. В обзоре ранних исследований по этому вопросу Кинг (King, 1958) отметил, что получаемые результаты зависели от возраста в период экспериментального воздействия, длительности этого периода, характера и интенсивности воздействия, возраста при тестировании и типа теста. У разных видов животных (обычно лабораторных крыс и домовых мышей) часто обнаруживался разный характер влияний. Нас не удивит, что и в пределах одного вида существует множество взаимодействий между генотипом и средой (Henderson, 1968a). Все это означает, что здесь трудно делать широкие обобщения. Мы должны учитывать все существенные факторы. Как мы уже видели в главе 7 при рассмотрении наследственности и психологических законов, природа организмов составляет важную часть всех законов. Эти сложности следует рассматривать скорее как интереснейшую проблему для дальнейшего изучения, а не как помеху в работе (Henderson, 1968a). Рассмотрим несколько важнейших факторов и некоторые результаты исследований в этой области. Экспериментальные воздействия. Во многих экспериментах ранние воздействия кажутся очень слабыми Можно в первые несколько дней жизни просто ежедневно забирать детеныша у матери и помещать его на несколько минут в жестяную банку. В других опытах на детеныша можно или воздействовать током, или систематически брать в руки и «ласкать», или делать что-нибудь еще во время изоляции от матери (рентгеновское облучение, введение веществ, вызывающих судороги, и т. п.). Разнообразные раздражители можно предъявлять, не забирая детеныша от матери. Чаще всего изучали влияние температурного стресса, но применялись также слуховой и другие типы стрессов. После прекращения вскармливания молоком детенышей можно выращивать в самых разнообразных условиях различной сложности. Влияние обогащения среды в раннем возрасте. Было описано множество эффектов, вызываемых обогащением окружающей среды в раннем возрасте Мы рассмотрим пять наиболее часто наблюдаемых явлений: 1) ускорение роста и развития; 2) ослабление эмоциональной реактивности; 3) улучшение результатов в задаче с научением; 4) более адекватные реакции организма на стресс; 5) изменения в развитии нервной системы (см. Daly, 1973). Часто оказывалось, что взятие детенышей в руки, удары тока и другие процедуры, проводимые в раннем возрасте, вызывают увеличение веса детенышей к моменту прекращения материнского вскармливания (см, например, Denenberg, Karas, 1961; Levine, Otis, 1958). Однако во многих работах таких эффектов получено не было (см. Daly, 1973). Сообщалось, что взятие в руки (handling) в раннем возрасте или удары тока влияют на «эмоциональность». Об изменениях в эмоциональности часто судят по поведению животных в «открытом поле». Хотя интерпретации, связанные с понятием эмоциональности, очень спорны, остается верным то, что, по многочисленным данным, в результате обогащения ранней среды происходят изменения частоты дефекации в открытом поле, уровне активности в том же тесте или обоих показателей (см, например, Denenberg, Smith, 1963, Denenberg et al, 1962) Удары тока или взятие в руки в раннем возрасте приводят к сходным изменениям. Оба этих воздействия обычно либо повышают активность, либо снижают частоту дефекации, либо вызывают и тот и другой эффект. Было неоднократно показано, что обогащение среды улучшает обучение целому ряду задач, особенно связанных с избеганием (см, например, Levine, 1956) Причина этого не всегда понятна. Возможно, что влияние обогащения внешней среды в период материнского вскармливания на научение вторична по отношению к другим изменениям, таким, как, например, изменения эмоциональных реакций в аппарате для тестирования Ряд работ показал, что после взятия в руки в раннем возрасте у крыс наблюдаются более адекватные реакции на стресс по сравнению с контрольными животными. Например, при помещении в новую обстановку они реагируют умеренным выбросом кортикостероидов, хотя в ситуациях с физической угрозой у них возникает более интенсивная реакция надпочечников, чем у контрольных животных (Levine, Mullins, 1966). Воздействия в раннем возрасте могут влиять на биохимию и анатомию мозга Тэпп и Марковиц (Тарр, Markowitz, 1963) сравнивали мозг крыс, подвергавшихся стимуляции, и крыс, которых в первые десять дней жизни никаким воздействиям не подвергали. У животных, которых брали в руки, вес подкорковых отделов мозга был больше, а уровень ацетилхолина в этих отделах – ниже. У крыс, выращенных в усложненной среде, отмечалось изменение ряда мозговых показателей, в том числе увеличение веса коры, повышение активности ацетилхолинэстеразы, увеличение толщины коры и более сильное ветвление дендритов нервных клеток (Rosenzweig et al, 1968; Greenough, 1975). Влияние времени воздействия. Почти все, кто изучает эффекты раннего опыта, согласны в том, что большое влияние на результаты оказывает время, когда производилось данное воздействие Многие из эффектов взятия в руки проявляются после осуществления этой процедуры в первые дни жизни, но не проявляются, если такому воздействию животных подвергают позднее. Обогащение внешней среды в пре- и постнатальный периоды может по-разному влиять как на биохимию и анатомию мозга, так и на поведение Хотя ясно, что возраст – важная переменная и что первые 5 дней жизни – это у мышей и крыс период особенно высокой чувствительности ко многим воздействиям, некоторые эффекты были обнаружены и при стимуляции в более поздние сроки. Поэтому Дененберг (Denenberg, 1968) пришел к выводу, что для эффектов обогащения внешней среды в раннем возрасте истинного критического периода не существует. Можно определить лишь период максимальной чувствительности к качеству и количеству стимуляции, к зависимым от нее переменным и другим подобным факторам Механизмы, лежащие в основе эффектов обогащения среды. Относительно этих механизмов было выдвинуто множество предположений (Russel, 1971; Daly, 1973). Мы рассмотрим три гипотетических фактора: 1) изменения температуры, 2) изменения поведения матери и 3) стресс в раннем возрасте. Согласно «гипотермической» гипотезе, как ее формулировал Шефер (Schaefer, 1968), влияние раннего опыта обусловлено изменением температуры Когда животное вынимают из гнезда и берут в руки, температура его тела изменяется. Для более поздних эффектов это изменение температуры может быть более важным, чем прямое влияние на нервную систему. Сходные эффекты были получены, когда животных подвергали только холодовому стрессу (см. Schaefer, 1968). Шефер считает, что нервная система новорожденного детеныша не настолько развита, чтобы реагировать на какое-либо прямое воздействие ранней стимуляции, и это делает роль температуры весьма правдоподобной. Второе объяснение состоит в том, что эффект раннего обогащения среды – результат изменений в поведении матери. Взятие в руки детенышей вызывает изменение в материнском поведении самки, в частности усиленное вылизывание детенышей (Priestnall, 1973; Lee, Williams, 1974). Животные, подвергающиеся многим воздействиям, издают особенно интенсивные ультразвуки, которые привлекают повышенное внимание матери. Таким образом, изменения в поведении могут быть в большей мере обусловлены усиленным вниманием со стороны матери, чем прямым воздействием на детеныша (Richards, 1966; Barnett, Walker, 1974; Thoman, Levin, 1970). Третья точка зрения исходит из наблюдений, касающихся стресса. Мы уже отмечали, что животные, которых в раннем возрасте брали в руки, более «адекватно» реагируют на умеренный и сильный стресс, нежели контрольные особи. Регуляция уровня гормонов осуществляется, по-видимому, с помощью отрицательной обратной связи через гипоталамус и гипофиз (см. гл. 12). При повышении уровня гормонов активность центров, управляющих секрецией гормонов, снижается. Эта система отрицательных обратных связей, напоминающая термостат, регулирующий температуру воздуха в помещении, была названа «гормоностатом» (Le-vine, Mullins, 1966). Предполагается, что в результате повышения активности надпочечников, вызванного стимуляцией в раннем возрасте, какие-то характеристики гормоностата изменяются так, что у взрослого животного он действует более эффективно. Эти три гипотезы, а также другие объяснения эффектов раннего опыта не исключают друг друга. В определенных случаях могут действовать все эти факторы, играя различную относительную роль в зависимости от ситуации. Влияние, осуществляемое через мать. Мы уже видели, что воздействие на детенышей влияет на поведение матери и что изменения материнского поведения могут служить важным посредником в механизме многих эффектов от обогащения внешней среды в раннем возрасте. Следующий вопрос касается возможных изменений в поведении детенышей, связанных с теми воздействиями, которые претерпевали их матери, когда в свою очередь были детенышами. Дененберг и Уимби (Denenberg, Whimbey, 1963) показали, что потомство матерей, которых брали в раннем возрасте в руки, отличалось от потомства контрольных животных и по весу тела к концу материнского вскармливания, и по показателям «открытого поля». Дененберг и Розенберг (Denenberg, Rosenberg„ 1967) сделали еще один шаг дальше и показали, что такое воздействие на крыс-самок влияет на активность и вес тела при отъеме от матери у их внуков. Суммарный эффект определялся взаимодействием ряда переменных. Интерпретация. Все согласны в том, что обогащение внешней среды в раннем возрасте глубоко влияет на поведение взрослого животного. Относительно природы переменных, вызывающих такие изменения, тоже нет больших разногласий. Однако эти переменные, по-видимому, взаимодействуют в различных условиях по-разному, и это приводит к невероятному разнообразию конкретных результатов и порождает много споров относительно их. интерпретации. Нужно сделать еще одно предостережение. Обычно принято Считать влияние раннего опыта «благоприятным», а условия содержания в лаборатории – «обедненными». Как подчеркнул Дэй-ли (Daly, 1973), такие обобщения следует делать с большой осторожностью. Мы не можем уверенно сопоставлять условия большинства лабораторных тестов с факторами, существенными для-выживания и успешного размножения животных в их природной среде. Поэтому трудно что-либо заключить о том, что окажется благоприятным для них в естественных условиях. Такая же осторожность нужна при оценке условий содержания животных в лаборатории. Большинство грызунов живет в норах. Эти норы устроены так, что колебания температуры в обычных лабораторных условиях могут быть значительно большими, чем в норе. Весьма вероятно, что лабораторная клетка создает для детенышей-сосунков более стимулирующее сенсорное окружение, чем нора. Дэйли (Daly, 1973, р. 435) предполагает даже, что «условия содержания в лаборатории оказываются для очень маленьких детенышей чрезмерно стимулирующими». Обеднение среды, (условия депривации). Очевидным дополнением к экспериментам с обогащением внешней среды служат опыты с ее обеднением. В типичном случае детенышей выращивают & условиях, когда они либо изолированы от особей своего вида, либо лишены каких-либо раздражителей. Многие из таких исследований были специально посвящены изучению онтогенеза перцептивных систем и будут обсуждаться в главе 10; здесь же мы рассмотрим влияние обедненной среды на эмоциональность и на общественное и репродуктивное поведение. Собаки. Томпсон и Хероп (Thompson, Heron, 1954) нашли, что шотландские терьеры, выращенные в условиях с рядом ограничений, решали задачи хуже, чем собаки, выросшие в домашних условиях. Мельцак (Melzak, 1968) суммировал результаты нескольких исследований, в которых собак выращивали в условиях с резко ограниченным сенсорным входом. После снятия этих ограничений такие собаки обнаруживали: 1) чрезвычайно высокий уровень активности, 2) аномальное восприятие зрительных, слуховых, обонятельных и болевых стимулов и 3) трудности в торможении неадекватных реакций. Например, они с трудом научались тормозить реакцию приближения к зажженной спичке. Как показал Фуллер (Fuller, 1967), изоляция в раннем возрасте сходным образом влияет на поведение животных во взрослом состоянии. Он предложил три альтернативных объяснения этих эффектов. Первое было связано с представлением о том, что внешние стимулы необходимы для развития нормальной организации нервной системы; тогда аномальное поведение – это результат недостаточного созревания нервной системы, для которого нужны индуцирующие сенсорные воздействия. Второе объяснение может состоять в том, что нервная система при рождении уже организована, но из-за отсутствия сенсорного входа претерпевает своего рода дегенерацию. В качестве третьей альтернативы Фуллер предположил, что, когда содержавшееся в изоляции животное впервые попадает в нормальную обстановку, воздействие неожиданного потока сенсорной информации может само по себе вызывать аномальное поведение. С этой точки зрения влияние сенсорной депривации связано не столько с нею самой, сколько с резким переходом от обедненной среды к нормальной. Фуллер показал, что ежедневного 10-минутного пребывания собаки в нормальных условиях в течение двух недель оказалось достаточно, чтобы устранить многие из эффектов депривации. Особое обращение с животными и применение транквилизаторов во время выхода из условий депривации оказывали действие, противоположное эффекту депривации. Макаки-резусы. После работ по импринтингу наиболее известными экспериментами с изменением раннего опыта, по-видимому, являются работы Гарри Ф. Харлоу и его сотрудников на макаках-резусах (например, Harlow, 1962, 1971; Harlow, Harlow, Suomi, 1971, Mason, 1960). Эти исследователи выращивали обезьян в условиях разной степени депривации. Многие были выращены в полной изоляции от других особей. В таких опытах использовали модельных «матерей», покрытых плюшем. Такие «матери» были необходимы многим детенышам как предмет, к которому можно было прижиматься и проявлять другие реакции, выражающие привязанность и в норме адресованные матери. Когда, однако, выращенные в условиях депривации обезьяны достигали зрелости, у них обнаруживались сильные нарушения репродуктивного поведения. Некоторые обезьяны не только отказывались спариваться, но и в более раннем возрасте обнаруживали склонность к аномальным манипуляциям с ротовой полостью, сжимали руками собственное тело, раскачивались, обнаруживали «апатию и безразличие к внешней стимуляции» (Harlow, Harlow, Suomi, 1971, р. 543). С большим трудом удалось добиться спаривания нескольких самок. Оказалось, что эти «матери без матерей» совершенно не способны выполнять свои функции: они игнорировали детенышей, жевали их ладони и ступни, бросали их лицом на пол. Если выросшую в изоляции обезьяну помещали с нормальными сверстниками, она навлекала на себя сильные проявления агрессии. Однако контакт с обезьянами более молодого возраста оказывал «лечебное» действие. Грызуны. В опытах с ранней депривацией, проводимых на грызунах, изучали главным образом последующее репродуктивное поведение. Хотя ранние исследования как будто говорили о другом, нам теперь ясно, что у выращенных в изоляции крыс и морских свинок обнаруживаются нарушения в акте спаривания (Valen-stein et al., 1955; Gerall, Ward, Gerall, 1967; Gruendel, Arnold, 1969). Обычно у них наблюдаются аномальные движения и отсутствие правильно ориентированной садки, так что копуляция оказывается неэффективной. Хотя у выращенных в изоляции самок не было видимых дефектов в материнском поведении, у их потомства обнаруживалась повышенная смертность, а также задержка в сроках открывания глаз по сравнению с потомством нормальных самок (Thoman, Arnold, 1968). КРАТКИЕ ВЫВОДЫ Онтогенетические изменения поведения можно проследить в течение всей жизни животного. В эмбриональном развитии появлению вызванных движений предшествует спонтанная подвижность. Постнатальное созревание связано с более медленными и постепенными изменениями в разнообразных формах поведения, включая сон, рефлексы, двигательную активность и общественное поведение. Импринтинг в форме «запечатления привязанности» могут вызывать весьма различные стимул-объекты, и он наиболее эффективен в довольно короткий чувствительный период. Эффект запечатления привязанности, по-видимому, обратим и может быть снят в результате предпочтения определенных звуковых сигналов, которое, видимо, бывает врожденным. Для полового импринтинга чувствительный период более длителен и наступает позднее, но этот вид импринтинга во многих случаях не так легко обратим. Обогащение среды и депривация в раннем возрасте сильно сказываются на поведении животных в более позднем возрасте. Специфические эффекты, получаемые при ранних средовых воздействиях, варьируют в зависимости от таких переменных, как возраст во время воздействия и во время тестирования, характер воздействия и природа зависимой переменной.
|
Последнее изменение этой страницы: 2019-04-09; Просмотров: 918; Нарушение авторского права страницы

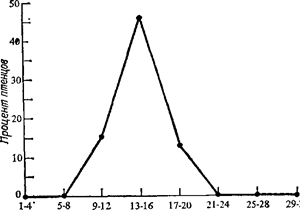 Рис 85 Чувствительный период для импринтинга. (Hess, 1959)
Рис 85 Чувствительный период для импринтинга. (Hess, 1959)