
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Моносахариды могут связываться друг с другом.Стр 1 из 12Следующая ⇒
Классификация углеводов. Углеводы, в первую очередь целлюлоза, являются самыми распространёнными органическими соединениями на Земле. В организме млекопитающих на долю углеводов приходится мерее 1% массы тела, однако их роль чрезвычайно велика. Углеводы, будучи компонентом протеогликанов, входят в состав соединительной ткани: углеводсодержащие белки глико- и мукопротеины являются составной частью защитных слизей организма6 углеводы входят в состав транспортных белков плазмы крови и иммунологически активных соединений: они формируют гликокаликс клеток. Углеводы являются основным источником энергии. По величине молекулярной массы углеводы делят на: · мносахариды · олигосахариды (2-10 моносахаридов) · полисахариды (более 10 моносахаридов) Моносахарид – это альдегид или кетон многоатомного алифатического спирта. Простейшими моносахаридами являются триозы: глицериновый альдегид (альдоза) и диоксиацетон (кетоза): Н Н │ │ С=О Н – С – ОН
Н – С – ОН С=О │ │ СН2ОН СН2ОН d-глицериновый диоксиацетон альдегид
Моносахариды с 4-мя углеродными атомами относятся к эритрозам, с 5-ю – к пентозам, с 6-ю – к гексозам, с 7-ю – к седогептулозам, с 8-ю – к октулозам. Моносахариды – оптически активные соединения. Их оптическая активность обуславливается асимметричным углеродным атомом (т.е. таким, у которого все 4 валентности связаны с разными радикалами). Таким асимметричным атомом уже обладает самая простейшая альдоза – глицериновый альдегид. Возможны 2 его пространственных варианта, являющихся зеркальным отражением друг друга, которые нельзя совместить при вращении. Их называют пространственными изомерами, или стереоизомерами; при этом стереоизомеры, в проекционной формуле которых ОН-группы расположены справа, называются D-формами (dexter – правый), если ОН-группы расположены слева, - то это L-форма (leavus – левый). Химические свойства у стереоизомеров одинаковы, но оптическая активность (угол вращения плоскости поляризованного света при прохождении его через раствор сахара) различна, т.е. D- и L-формы вращают плоскость поляризованного луча на одну величину, но в противоположном направлении. Экспериментально установлено, что для глицеринового альдегида правовращающей (+) является D-форма. Вращение влево обозначается знаком – При удлинении углеродной цепи в моносахаридах число асимметричных атомов углерода увеличивается, при этом количество стереоизомеров будет составлять 2n (n – количество асимметричных атомов С). Так, у гексоз с 4 асимметричными углеродными атомами будет 16 стереоизомеров и 8 различных химически отличающихся соединений. Природа предпочитает стереоизомеры, у которых последний асимметричный атом углерода имеет D-форму, поэтому их называют D-моносахаридами. Ферменты клеток строго различают стереоизомеры, синтезируя и вызывая распад исключительно D-моносахаридов.
Рис.1.Химические свойства различных моносахаридов схожи в силу сходства их строения:
1.Углеводы обладают свойствами восстановителей (благодаря наличию альдегидной группы в составе их молекулы), что даёт возможность проводить качественное и количественное определение сахаров. На этом свойстве базируется о-толуидиновый метод определения уровня глюкозы в крови и реакции (Троммера, Ниландера) определения сахара в моче. Однако эти методы недостаточно специфичны, так как помимо глюкозы цветную реакцию дают и другие редуцирующие сахара. 2..При окислении моносахаридов образуются уроновые кислоты, из которых важнейшей является глюкуроновая кислота, входящая в состав основного вещества соединительной ткани. 3..Моносахариды способны образовывать эфиры, особо важны фосфорные эфиры гексоз (глюкозы, фруктозы, галактозы) и пентоз (рибозы и дезоксирибозы), так как именно фосфорилированные сахара участвуют в реакциях метаболизма. 4..Моносахариды могут присоединять аминогруппу (образуются глюкозамины) и ацетилироваться. Фосфофрукто-
Альдолаза В альдолаза А Глицериновый диоксиацетон Альдегид фосфат
Сорбитолдегидрогеназа
Обмен галактозы. Галактоза входит в состав молочного сахара лактозы. В печени галактоза фосфорилируется галактокиназой с образованием галактозо-1-фосфата. Следующая реакция катализируется уридилтрансферазой, переносящей УДФ от УДФ-глюкозы на галактозо-1-фосфат. Наконец, УДФ-галактоза эпимеризуется ( эпимераза ) в УДФ-глюкозу, которая может превращаться в глюкозо-1-фосфат ферментом пирофосфорилазой. галактокиназа АТФ
Уридилтрансфераза Эпимераза Гликогенфосфорилаза
Н3РО4
Места ветвления разъединяет «деветвящий фермент» - амило-(1→ 6)-гликозидаза – рис. 12.
Рис.12. Функционирование деветвящего фермента.
Вначале этот фермент переносит 3 остатка глюкозы на другую ветвь гликогена (трансферазная активность фермента), затем он гидролизует α (1→ 6)гликозидную связь и освобождает молекулу глюкозы у точки ветвления (глюкозидазная активность фермента). Затем, после действия этого фермента, включается вновь гликогенфосфорилаза. Отщепившийся под влиянием гликогенфосфорилазы глюкозо-1-фосфат превращается в глюкозо-6-фосфат (фермент – фосфоглюкомутаза):
Фосфоглюкомутаза
В мышцах, в отличие от печени, глюкозо-6-фосфат не превращается в свободную глюкозу, так как в миоцитах отсутствует фермент глюкозо-6-фосфатаза. В печени (а также в почках) под действием этого фермента образуется глюкоза: Фосфатаза глюкозо-6-фосфата
Только свободная глюкоза, в отличие от глюкозо-6-фосфата, способна проходить через плазматическую мембрану клеток в межклеточное пространство. Таким образом, печень является органом, поддерживающим нормальный уровень глюкозы в крови. В этом заключается её огромная роль в поддержании жизнедеятельности организма. Главным регулируемым ферментом гликогенолиза является гликогенфосфорилаза. Фосфорилаза может находиться либо в неактивном, дефосфорилированном, состоянии – фосфорилаза b, либо в активном, фосфорилированном, состоянии – фосфорилаза a. Неактивная форма фермента, состоящая из 2-х субъединиц, превращается в активную, состоящую из 4-х субъединиц, с помощью киназы фосфорилазы b. киназа фосфорилазы b
фосфатаза Киназа фосфорилазы b может также находится в активной и неактивной формах. Превращение неактивной киназы в активную осуществляется цАМФ-зависимым ферментом протеинкиназой: Протеинкиназа Неактивная Активная
Содержание цАМФ в гепатоцитах подчиняется гормональному контролю, в котором принимают участие инсулин, адреналин и глюкагон (рис 13).
Рис.13 Аденилатциклазный механизм. Адреналин и глюкагон, активируя аденилатциклазу, способствуют образованию цАМФ, который запускает «каскадный механизм» фосфорилирования ферментов распада и синтеза гликогена. В результате фосфорилирования образуется фосфорилированная, т.е. активная, гликогенфосфорилаза и фосфорилированная, т.е. неактивная гликогенсинтаза. В этих условиях будет осуществляться распад гликогена. Напротив, под действием инсулина, включающего механизм де фосфорилирования ключевых ферментов, появятся де фосфорилированная, т.е. не активная гликогенфосфорилаза, и де фосфорилированная, т.е. активная, гликогенсинтаза. В этих условиях будет происходить синтез гликогена. Активность обоих обоих ключевых ферментов синтеза и распада гликогена в печени регулируется также Са++ – выход кальция из внутриклеточных депо в цитозоль клетки контролируется инозитолтрифосфатом (ИТФ), который освобождается фосфолипазой С при распаде входящего в состав мембраны фосфатидилинозитол пирофосфата (ФИФФ). Заключение. Т.о. глюкоза, поступившая в клетки с помощью транспортных систем приобретает заряд (-2) и начинается дальнейшее окисление эфира- глюкозо-6 фосфата. Метаболизм глюкозо-6 фосфата включает следующие направления: 1. Дихотомический путь протекает как в анаэробных (гликолиз), так и аэробных условиях (в этом пути «туловище» глюкозы расщепляется пополам – дихотомия); 2. Апотомическом пути (apex - верхушка). Этот путь называется также пентозофосфатным. 3. Синтез гликогена как резервного материала клеток 4. Окисление глюкозы и последующее отщепление 6-го углеродного атома глюкозы – глюкуроновый путь, с последующим образованием аминосахаров- глюкозаминогликанов( ГАГ). доц.кафедры
Министерство здравоохранения Республики Беларусь Учреждение образования «Гомельский государственный медицинский университет» Кафедра биологической химии Обсуждено на заседании кафедры (МК или ЦУНМС)____________________ Протокол № _______
ЛЕКЦИЯ
По биологической химии
для студентов 2-го курса лечебного факультета Тема: Углеводы 2. Тканевой обмен углеводов Время 90 мин
Учебная цель: Сформировать представления о путях и механизмах обмена глюкозы в организме
ЛИТЕРАТУРА 1.Биохимия человека:, Р.Марри, Д.Греннер, П.Мейес, В.Родуэлл.- М.книга, 2004.- т.1.- с. 172-177., 181-188., 191-194. 2.Основы биохимии: А.Уайт, Ф.Хендлер, Э.Смит, Р.Хилл, И.Леман.-М. книга, 1981, т. 2,.с.612-642. 3.Наглядная биохимия: Кольман., Рем К.-Г-М.книга 2004г., -с. 138-140., 148-152. 4. Биохимические основы...под. ред. член- корр. РАН Е.С. Северина. М.Медицина, 2000.-с.98-111.
МАТЕРИАЛЬНОЕ ОБЕСПЕЧЕНИЕ 1.Мультимедийная презентация
РАСЧЕТ УЧЕБНОГО ВРЕМЕНИ
Всего 90 минут
HСОH │ СН2― О― РО3Н2
Фосфоглицериновый Альдегид (3-ФГА)
4-я реакция обратима. Катализирующий её фермент называется альдолазой, так как расщепляется альдол – фруктозо-1, 6-дифосфат. Равновесие реакции сдвинуто в сторону распада фруктозо-1, 6-дифосфата, поскольку образующийся 3-ФГА расходуется в дальнейших реакциях гликолиза. Именно в силу последнего обратимая триозофосфатизомеразная реакция также смещается в сторону образования 3-ФГА. Определение активности альдолазы используют в энзимодиагностике при заболеваниях, связанных с повреждением или гибелью клеток: так, при остром гепатите активность этого фермента может увеличиваться в 5-20 раз, при инфаркте миокарда – в 3-10 раз, при миодистрофии – в 4-10 раз. Таким образом, в подготовительной стадии гликолиза фактически образуется 2 молекулы 3-ФГА. Следующая стадия гликолиза – гликолитическая оксидоредукция – является более сложной. 5-я реакция. В этой реакции 3-ФГА окисляется дегидрогеназой 3-фосфоглице-ринового альдегида. Фермент состоит из 4-х одинаковых субъединиц, в состав его активного центра входит SH-группа. Дегидрогеназа 3-ФГА нуждается также в коферменте НАД+. Фермент окисляет свой субстрат (3-ФГА), перенося водород альдегидной группы 3-фосфоглицеринового альдегида на кофермент НАД+, который при этом восстанавливается в НАДН∙ Н+ в присутствии неорганического фосфата. Выделяющейся при окислении 3-ФГА энергии достаточно для образования высокоэргического (т.е. заключающего в себе м а к р о э р г и ч е с к у ю связь) соединения – 1, 3-дифосфоглицериновой кислоты (1, 3-ДФГК). Образование НАДН∙ Н+ из окисленной формы этого кофермента осуществляется в ходе реакции связывания 3-ФГА с SH-группой активного центра фермента – дегидрогеназы 3-фосфоглицеринового альдегида: CH2O℗ CH2O℗ CH2O℗ CH2O℗
CHOH CHOH CHOH CHOH │ 1 │ 2 │ 3 │ H― C═ O H― C― OH C═ O C═ O 3-ФГА │ │ │ S S O
ДФГК SH
6-я реакция. Эта реакция получила название реакции субстратного фосфорилирования – фосфорилирование АДФ за счёт энергии макроэргического субстрата. Реакция сопровождается выделением значительного количества свободной энергии, поэтому её равновесие сдвину вправо. Она может стать обратимой при избытке 3-фосфоглицерата.
СН2О― РО3Н2 Mg2+ СН2О― РО3Н2 | фосфоглицераткиназа |
| |
О ~ РО3Н2 ОН
ДФКГ 3-фосфоглицерат Я реакция. Изомеризация 3-фосфоглицерата в 2-фосфоглицерат:
СН2О― РО3Н2 СООН | фосфоглицеромутаза |
| |
ОН Фосфоенолопируват ПВК Реакция образования ПВК (пировиноградной кислоты) необратима, так как протекает со значительным падением свободной энергиию. 10-я реакция. Заключительная реакция гликолиза – образование лактата. Реакция обратима. Молочная кислота образуется при восстановлении пирувата ферментом лактатдегидрогеназой (ЛДГ), коферментом которого является НАДН∙ Н+ (образование этого кофермента см. в 5 –й реакции).
СООН СН3 | | С═ O + НАДН∙ Н+ Лактатдегидрогеназа Н― С― ОН + НАД+
СН3 СН3 ПВК Лактат
Итак, гликолиз завершается образованием лактата. В мышцах молочная кислота не используется – она поступает с током крови в печень, где вновь превращается (благодаря обратимости ЛДГ-азной реакции) в пируват. Изоферменты лактатдегидрогеназы принимают участие в контроле гликолиза: так, в сердечной мышце преобладает ЛДГ1, который ингибируется даже небольшими концентрациями пирувата, что затрудняет образование молочной кислоты в кардиомиоцитах и способствует дальнейшему окислению (а не восстановлению) пирувата; в скелетных мышцах преобладает изофермент ЛДГ5, активно превращающий ПВК в лактат в анаэробных условиях. Гликолиз протекает в цитоплазме клетки, он не нуждается в участии кислорода для получения клеткой энергии. В ходе гликолиза в 2-х реакциях субстратного фосфорилирования (реакции 6-я и 9-я) образуется 4 молекулы АТФ --(напоминаем, что в реакцию вступают 2 триозы), однако в подготовительной стадии 2 молекулы АТФ расходуются (реакции 1-я и 3-я), таким образом, полезный энергетический выход гликолиза составляет 2 молекулы АТФ. Мебрана митохондрий непроницаема для образующегося оксалацетата (ЩУК). Однако, превращаясь в яблочную кислоту при участии митохондриальной НАД+-зависимой малатдегидрогеназы, малат легко покидает митохондрию и в цитозоле клетки окисляется в ЩУК при участии цитоплазматической НАД+-зависимой малатдегидрогеназы. Дальнейшее превращение ЩУК в фосфоенолПВК происходит в цитозоле клетки.
Рис. 1.Спиртовое брожение глюкозы
Как видно из Рис. 1.образование ацетальдегида из ПВК осуществляется путём прямого декарбоксилирования пирувата. Реакция катализируется пируват-карбоксилазой сучастием её кофермента – тиаминпирофосфата. Уксусный альдегид – чрезвычайно токсичное соединение. Он обезвреживается алкогольдегидрогеназой, коферментом которой является НАДН ·Н+.Экзогенный этанол в печени окисляется в ацетальдегид с помощью той же алкогольдегидрогеназы (кофермент – НАД+). Именно за счёт накопления ацеальдегида при потреблении спиртных напитков и происходит отравление организма. Обезвреживание этого соединения осуществляется альдегиддегидрогеназой, активность которой (а значит и переносимость алкоголя) значительно варьирует у разных лиц и наций. Альдегиддегидрогеназа превращает ацетальдегид в уксусную кислоту. Последняя активируется, превращаясь в ацетил-КоА, который «сгорает» в цикле трикарбоновых кислот с образованием энергии. В обезвреживании этанола принимают также участие микросомная система детоксикации гепатоцитов и фермент каталаза. Однако их роль менее значима. Систематическое потребление алкоголя приводит к циррозу печени и увеличивает риск развития рака (причём не только печени), особенно на фоне хронического воздействия малых доз облучения. Цитрат Изоцитрат
COOH α -кетоглутарат Сукцинил-КоА Cукцинил-КоА Сукцинат
нуклеозидфосфаткиназа
Сукцинат Фумарат
7 реакция – образование яблочной кислоты ферментом фумаразой. Фумараза (фумаратгидратаза) гидратирует фумаровую кислоту – при этом образуется яблочная кислота, причём её L-форма, так как фермент обладает стереоспецифичностью.
СООН СООН │ Н2О │
СН фумараза Н–С – Н │ │ СООН СООН Фумарат L-Малат
8 реакция – образование оксалацетата. Реакция катализируется малатдегидрогеназой, коферментом которой служит НАД+. Образовавшийся под действием фермента оксалацетат вновь включается в цикл Кребса и весь циклический процесс повторяется.
СООН СООН │ │ Н О–С– Н НАД+ НАД Н ∙ Н+ С=О
CH2 CH2 │ │ СООН СООН Малат ЩУК
Последние 3 реакции обратимы, но поскольку НАДН∙ Н+ захватывается дыхательной цепью, равновесие реакции сдвигается вправо, т.е. в сторону образования оксалацетата. Как видно, за один оборот цикла происходит полное окисление, «сгорание», молекулы ацетил-КоА. В ходе цикла образуются восстановленные формы никотинамидных и флавиновых коферментов, которые окисляются в дыхательной цепи митохондрий. Таким образом, цикл Кребса находится в тесной взаимосвязи с процессом клеточного дыхания. Пируваткарбоксилаза
Глутатион –S-S-глутатион
НАДФН∙ Н+ Н2О Глутатионредуктаза Глутатионпероксидаза НАДФ+ Н2О2 Глутатион-SH МетНb Hb Витамин С
Рис.2 Значение ПФП Недостаточность глюкозо-6-фосфатдегидрогеназы является наиболее частым дефектом ферментативных систем эритроцита. При низкой активности этого фермента возникает дефицит НАДФН∙ Н+, что ограничивает функцию глутатионредуктазы, т.е. нарушается восстановление глутатиона. Падение концентрации глутатиона-SH влечёт за собой снижение активности глутатионпероксидазы. Это приводит к накоплению продуктов ПОЛ, следствием чего являются метгемоглобинемия и, поскольку избыток перекисных продуктов способен разрушить эритроцитарную мембрану, – гемолитическая анемия Надо заметить, что НАДФН∙ Н+, в отличие от НАДН∙ Н+, не участвует в окислительном фосфорилировании, протекающем в митохондриях, и не служит, таким образом, для получения энергии. Велико значение пентозофосфатного пути как поставщика рибозы-5-фосфата, необходимого для построения мононуклеотидов (АМФ, АДФ, АТФ, ГМФ и т.д.), олигонуклеотидов, коферментов (ФМН, ФАД, НАД, НАДФ), нуклеиновых кислот. Апотомический путь тесно связан с гликолизом. В зависимости от тех или иных условий оба пути могут переключаться друг на друга, поскольку у них имеются общие метаболиты (например, фруктозо-6-фосфат, 3-ФГА). Однако в отличие от гликолиза в пентозофосфатном пути используется другой кофермент (НАДФ+ вместо НАД+), в нём образуется СО2 (чего нет в гликолизе) и он не обладает энергетической функцией. Регуляция. Главными регулируемыми (ключевыми) ферментами пентозофосфатного пути являются 2 дегидрогеназы его окислительной части: глюкозо-6-фрсфатдегидрогеназа и дегидрогеназа 6-фосфоглюконата. Индукторами биосинтеза этих ферментов является инсулин. Активность дегидрогеназ увеличивается при поступлении углеводов в организм и снижается при голодании и диабете. Именно поэтому они считаются адаптивными ферментами
Глюкоза УДФ-глюкоза УДФ-глюкуроновая кислота Фосфофрукто-
Альдолаза В альдолаза А Глицериновый диоксиацетон Альдегид фосфат
Сорбитолдегидрогеназа
Обмен галактозы. Галактоза входит в состав молочного сахара лактозы. В печени галактоза фосфорилируется галактокиназой с образованием галактозо-1-фосфата. Следующая реакция катализируется уридилтрансферазой, переносящей УДФ от УДФ-глюкозы на галактозо-1-фосфат. Наконец, УДФ-галактоза эпимеризуется ( эпимераза ) в УДФ-глюкозу, которая может превращаться в глюкозо-1-фосфат ферментом пирофосфорилазой. галактокиназа АТФ
Уридилтрансфераза Эпимераза Рис.8 Метаболизм галактозы Недостаточность галактокиназы проявляется катарактой (галактитол – осмотически активное соединение, вызывающее помутнение хрусталика глаза). Наиболее распространённым и тяжёлым является врождённый дефект уридилтрансферазы (галактозо-1-фосфат-уридилтрансферазы). Он проявляется синдромом галактоземия. При этом заболевании из-за недостаточности уридилтрансферазы в крови резко повышается содержание галактозо-1-фосфата и галактозы, дающие положительную реакцию на «сахар» крови. Сахар обнаруживается в моче (галактозурия). Синдром галактоземии проявляется желтухой новорождённых, гепатомегалией, задержкой психического развития. Заподозрить этот дефект можно на основании рвоты, возникающей после кормления ребёнка грудью, поноса, прогрессирующей катаракты. При исключении из рациона галатозы (молока) проявления заболевания значительно уменьшаются, однако катаракта не исчезает. Гликогеновые болезни.
Гликогеновые болезни относятся к наследственным нарушениям обмена. Они делятся на две основных группы: 1. Гликогенозы – развиваются в результате недостаточной активности или отсутствия ферментов, ответственных за распад гликогена. 2. Агликогенозы – результат недостаточности ферментов синтеза гликогена. Фосфоглюкомутаза Глюкозо-1-фосфат
УДФ-глюкоза
VI тип - печёночная) Гликоген
ф-нт ветвле- γ -амилаза (лизосом) α (1→ 4) ния α (1→ 6) (II тип) (IV тип) ф-нт развет- вления α (1→ 6) мальтаза (лизосом)
Гликоген Рис. 9 Метаболизм гликогена 2 тип – отсутствие γ -амилазы . Отмечается генерализованное поражение всех органов. 3 тип – дефект фермента разветвления (амило-1, 6-гликозидазы). В печени и мышцах накапливается гликоген, молекула которого имеет очень длинные боковые ветви, так как она не может быть расщеплена в местах ветвления. 4 тип – дефект фермента ветвления. Структура молекулы гликогена представлена очень короткими ветвями. 5 тип (болезнь Мак Ардля) и 6 тип (болезнь Герше) – недостаточность фосфорилазы. Структура гликогена при этом не нарушена. 5 типу гликогенозов относят дефект фосфорилазы мышц, 6-й – печени. Соответственно, клиническая симптоматика 5 типа характеризуется резкой мышечной слабостью (из-за отсутствия распада гликогена страдает энергетика мышцы), 6 типа – гепатомегалией и накоплением гликогена в лейкоцитах. Имеются также и некоторые другие разновидности гликогенозов. Характерным для всех гликогенозов является гепатомегалия, мышечная слабость, гипогликемия натощак. Введение адреналина таким больным вызывает не гипергликемию, а гиперлактатацидемию. Жизнь таких больных укорачивается. При агликогенозах в результате нарушения синтеза гликогена страдают энергетические ресурсы клетки. Сахарный диабет тип I. Диабет- это выделение какого- либо метаболита с мочой. По распространенности занимает 3- место среди других заболеваний, после ССЗ и рака. В мире насчитывается100 млн чел больных СД, и каждые 10-15 лет число больных СД удваивается. Наиболее подвержены риску к заболеванию СД малообеспеченные лица. проживающие в индустриально развитых странах. Диабет 1 типа развивается в юношеском возрасте, иногда в детстве, и очень редко у взрослых. Протекает тяжелее, чем СД 2- типа. При отсутствии врачебного контроля- возможны острые осложнения. Встречается в 10 раз меньше, чем СД 2 типа. Регуляция углеводного обмена осуществляется гормонами Инсулином и Глюкагоном. Эти же гормоны влияют на механизм депонирования и мобилизации гликогена, а также на метаболизм жиров. Инсулин и Глюкагон это главные регуляторы изменений метаболизма при смене состояний пищеварения и голодания ( абсорбтивное и постабсорбтивное состояния). Пищеварение длится 10-15 час в сутки, а расход энергии все 24 час.( снижен ночью). Поэтому часть энергии депонируется, для того, чтобы использоваться в постабсорбтивный период. Печень, жировая ткань и мышцы- главные органы, при этих изменениях. При 3х разовом питании, смена режимов происходит 3 раза в сутки. Но эта смена выражена нестойко, т.к. в течение дня промежутки между приемами пищи небольшие (5-6 ч) и постабсорбтивный период едва- едва успевает начаться, как наступает время очередного приема пищи. Типичное постабсорбтив. состояние – это утро перед завтраком, после 10 часового перерыва в еде. Немецкий ученый- Эммануил Кант считал разумным есть 1 раз/ сутки. За сутки запас гликогена в печени заканчивается и единственным источником остается глюкоза, синтезируемая в ГНГ. В этот период глюкоза потребляется. Нервными клетками, а другие клетки получают Энергию за счет окисления ЖК., и кетовых тел, синтезируемых в печени.. Эта модель считается как постабсорбтивная или рассматривается как кратковременное голодание. Существуют два основных механизма переноса Гл в клетки-: активный транспорт, зависящий от градиента конц ионов Nа+, и облнегченная диффузия. Соответственно есть два типа рецепторов Дл Гл. Рецепторы, Зависимые от конц. Nа есть только в почках и кишечнике, они обеспечивают реабсорюцию Гл из почечных канальцев и всасывание ее из просвета кишечника против градиента конц. Рецепторы облегченной диффузии – есть во всех тканях. В тканях человека есть 5 разных Глют. ГЛЮТ 1- в плаценте, мозге, почках, толстой кишке, в бета Кл-ках островков Лангенгарса, меньше в жировой ткани и мышцах. ГЛЮТ 2 преимущественно в печени, энтероцитах, в проксимальных тубулярных Кл-х почек, в бета Кл-х Лангенгарса. ГЛЮТ 3 во многих тканях включая мозг, плаценту, почки. ГЛЮТ 4 – единственный переносчик, регулируемый инсулином, сод-ся в мыщцая ( скелетных и сердечной) и жировой ( инсулинзав.ткани) ГЛЮТ 5 – главный переносчик гл в базальном состоянии т.е. в отсутствии стимуляции Инсулина. Основные энергоносители- это ГЛ и ЖК. Регуляцию метболизма Углеводов ИНС и Глюкогоном нельзя рассматривать по отдельности В крови постоянно присутствуют оба гормона, но их соотношение их меняется. Оба действуют на одни и те же мишени. Гликоген через путь RAS однгвременно активирует гликогенсин-зу и ингибирует гликогенфосфорилазу, глюкагон через цАМФ зависимые протеинкиназы ингибирует гликогенсинтазу и активирует гликогенфосфорилазу.. ИНс подавляет не базальную скоростьГНГ, а только скорость стимулированную глюкагоном.. Кроме того ИНс снижает секрецию и самого глюкагона. ИНс образуется из препроинс в результате посттрансляционной модификации. Недавно было обнаружено, что С- пептид в физол. Конц. Стимулирует потребление гл. клетккми здорового человека и б-х СД втакой степени как и Инс. Глюкоза- регулирует экспрессию гена ИНС, а также др. белков, участвующих в обмене основных энергоносителей. В печени, под.Ж. и жировых Кл. при потреблении углеводов, происходит транскрипция целого ряда генов, связанных с метаболизмом углеводов. Инс участвует в регуляции таких клеточных процессов как метаболизм, трансмембранный перенос ионов, АмК, , Гл, синтез и распад белков, ИНС влияет на ядерные процессы- репликацию и транскрипцию, участвует в регуляции клеточной пролиферации, и дифференцировке, а также трансформации клеток. Гиперинсулинизм Фосфофрукто-
Альдолаза В альдолаза А Глицериновый диоксиацетон Альдегид фосфат
Сорбитолдегидрогеназа Обмен галактозы. Галактоза входит в состав молочного сахара лактозы. В печени галактоза фосфорилируется галактокиназой с образованием галактозо-1-фосфата. Следующая реакция катализируется уридилтрансферазой, переносящей УДФ от УДФ-глюкозы на галактозо-1-фосфат. Наконец, УДФ-галактоза эпимеризуется ( эпимераза ) в УДФ-глюкозу, которая может превращаться в глюкозо-1-фосфат ферментом пирофосфорилазой. галактокиназа АТФ
Уридилтрансфераза Эпимераза Гликогеновые болезни.
Гликогеновые болезни относятся к наследственным нарушениям обмена. Они делятся на две основных группы: 1. Гликогенозы – развиваются в результате недостаточной активности или отсутствия ферментов, ответственных за распад гликогена. 2. Агликогенозы – результат недостаточности ферментов синтеза гликогена. Фосфоглюкомутаза Глюкозо-1-фосфат
УДФ-глюкоза
VI тип - печёночная) Гликоген
ф-нт ветвле- γ -амилаза (лизосом) α (1→ 4) Популярное:
|
Последнее изменение этой страницы: 2016-03-17; Просмотров: 1578; Нарушение авторского права страницы

 │ │
│ │  Моносахариды могут иметь незамкнутые и циклические формы (5-членное – фуранозное кольцо, 6-членное – пиранозное кольцо). Образование кольца приводит к появлению асимметрии первого углеродного атома: ОН-группа может быть направлена вверх или вниз по отношению к кольцу (соответственно возникают α и β формы), находящиеся в равновесии с формой открытой цепи.
Моносахариды могут иметь незамкнутые и циклические формы (5-членное – фуранозное кольцо, 6-членное – пиранозное кольцо). Образование кольца приводит к появлению асимметрии первого углеродного атома: ОН-группа может быть направлена вверх или вниз по отношению к кольцу (соответственно возникают α и β формы), находящиеся в равновесии с формой открытой цепи.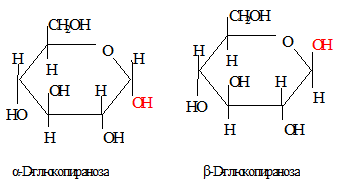
 фруктокиназа АТФ киназа АТФ
фруктокиназа АТФ киназа АТФ 
 фруктоза фруктозо1-фосфат фруктозо-1, 6-дифосфат
фруктоза фруктозо1-фосфат фруктозо-1, 6-дифосфат  АТФ
АТФ галактоза галактозо-1-фосфат УДФ-глюкоза
галактоза галактозо-1-фосфат УДФ-глюкоза  (ГЛИКОГЕН)n (ГЛИКОГЕН)n-1 + глюкозо-1-фосфат
(ГЛИКОГЕН)n (ГЛИКОГЕН)n-1 + глюкозо-1-фосфат
 Глюкозо-1-фосфат Глюкозо-6-фосфат
Глюкозо-1-фосфат Глюкозо-6-фосфат Глюкозо-6-фосфат Глюкоза + Н3РО4
Глюкозо-6-фосфат Глюкоза + Н3РО4
 2 фосфорилаза b + 4 АТФ фосфорилаза a + 4 АДФ
2 фосфорилаза b + 4 АТФ фосфорилаза a + 4 АДФ киназа фосфорилазы b киназа фосфорилазы b
киназа фосфорилазы b киназа фосфорилазы b
 │ │ НАД+ НАДН∙ Н+ │ │
│ │ НАД+ НАДН∙ Н+ │ │ 
 Н― С― ОН + АДФ АТФ + Н― С― ОН
Н― С― ОН + АДФ АТФ + Н― С― ОН
 С═ О С═ О
С═ О С═ О Н― С― ОН + АДФ АТФ + Н― С― О― РО3Н2
Н― С― ОН + АДФ АТФ + Н― С― О― РО3Н2 | |
| | ГТФ + АДФ ГДФ + АТФ
ГТФ + АДФ ГДФ + АТФ СН ОН –СН
СН ОН –СН ║ │
║ │  │ │
│ │  ПВК + НСО3- – биотин + Н2О ЩУК + АДФ + Н3РО4
ПВК + НСО3- – биотин + Н2О ЩУК + АДФ + Н3РО4 фруктокиназа АТФ киназа АТФ
фруктокиназа АТФ киназа АТФ 
 гликогенсинтаза фосфорилаза (V тип - мышечная,
гликогенсинтаза фосфорилаза (V тип - мышечная, 
 (III тип) мальтоза глюкоза
(III тип) мальтоза глюкоза