|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Ауксины. Химическая природа, биосинтез, физиологическая роль, практическое применение.Стр 1 из 10Следующая ⇒
Ауксины. Химическая природа, биосинтез, физиологическая роль, практическое применение.
Основным ауксином является индолил-3-уксусная кислота (ИУК). Обнаружены и другие природные вещества с ауксиновой активностью, такие как индолил-3-ацетонитрил, 4-хлор-3-индолилуксусная и фенилуксусная кислоты, однако их ауксиновая активность существенно слабее. ИУК синтезируется из триптофана в верхушках побега и перемещается сверху вниз по паренхимным клеткам со скоростью 10-15 мм/час благодаря особому механизму полярного транспорта. Возможно также более быстрое передвижение ауксинов по транспортным каналам растения. Ауксины обладают многообразным физиологическим действием и жизненно важны для роста и развития растений. Ауксины необходимы для деления и растяжения клеток, для формирования проводящих пучков и корней, способствуют разрастанию околоплодника. Ауксины обусловливают явление апикального доминирования, т. е. тормозящее действие апикальной почки на рост пазушных почек. Ауксины играют первостепенную роль в ростовых движениях: фото- и геотропизме и настиях. Ауксины усиливают аттрагирующее действие органов и тканей (т. е. их способность притягивать питательные вещества) и во многих случаях задерживают их старение. При реализации многих физиологических программ ауксины взаимодействуют с цитокининами и другими фитогормонами. При высоких концентрациях ауксины повышают образование своего антагониста -- фитогормона этилена. Первичное действие ауксинов направлено на изменение активности (активацию или репрессию) определенного набора компетентных генов, характерного для данной ткани. ИУК также активирует АТФазу плазмалеммы, вызывая выкачивание протонов из клетки и закисление клеточной стенки. Это приводит к размягчению матрикса стенки, что делает возможным рост клеток растяжением. На практике ауксины и их синтетические аналоги применяют для размножения клеток и растений в стерильной культуре и для получения трансгенных растений (совместно с цитокининами). Их часто используют для предотвращения предуборочного опадения плодов древесных культур, получения бессемянных плодов томатов, огурцов, баклажанов, перца и др., а также как стимуляторы корнеобразования у черенков; в высоких дозах - как гербициды и дефолианты (2, 4-дихлорфеноксиуксусная кислота и др.). Ауксины синтезируются многими фитопатогенными и симбиотическими микроорганизмами, что помогает последним воздействовать на клетки растения-хозяина. Аэробная дыхательная цепь. Окислительное фосфолирование. 2.
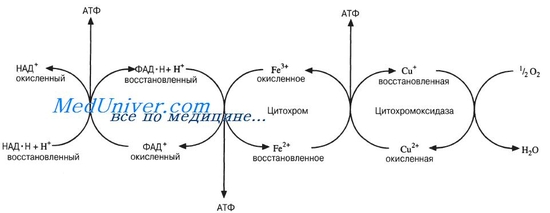
Схема дыхательной цепи.
В процессе окисления пировиноградной кислоты в цикле Кребса образовались пары водорода 2Н, которые мы можем рассматривать как 2Н+ + 2е. Именно в таком виде они, акцептированные НАД и ФАД, передаются по цепи переносчиков. В процессе переноса протонов и электронов важную роль играют ферменты, относящиеся к классу оксидоредуктаз. Оксидоредуктазы, участвующие в дыхательной цепи, делятся на следующие основные группы. Пиридиновые дегидрогеназы, у которых коферментом служит НАД или НАДФ, отнимают два протона и два электрона от субстрата. При этом к коферментам присоединяются один протон и два электрона. Протон и один электрон связываются с атомом углерода в молекуле НАД, а второй электрон нейтрализует положительный заряд атома азота. Один протон выделяется в среду. НАД+ и НАДН хорошо растворимы в воде и присутствуют в цитоплазме и митохондриях. Коферменты НАД и НАДФ связаны с ферментом с помощью ионов металла и сульфгидрильных группировок. В зависимости от белкового носителя, к которому присоединен кофермент (НАД или НАДФ), различают более 150 пиридиновых дегидрогеназ. Каждая из них специфична по отношению к определенному субстрату. Необходимо учитывать, что НАД и НАДФ могут воспринимать протоны и электроны лишь в том случае, если субстрат имеет более отрицательное значение потенциала по сравнению с ними. Окислительное фосфорилирование – процесс переноса неорганического фосфата с макроэргических соединений на АДФ и образование АТФ. Влияние внутренних и внешних факторов на проницаемость протоплазмы Ищите в тетрадях
Влияние водного режима, концентрации СО2 на интенсивность фотосинтеза.
Чем выше концентрация СО2, тем интенсивней идет процесс фотосинтеза.( При концентрации 0, 03 % интенсивность фотосинтеза составляет лишь 50 % от максимальной, которая достигается при 0, 3% СО2) Водный режим Вода непосредственно участвует в фотосинтезе как субстрат окисления и источник кислорода. Другой аспект влияния содержания воды на фотосинтез состоит в том, что величина оводненности листьев определяет степень открывания устьиц и, следовательно, поступления С02 в лист. При полном насыщении листа водой устьица закрываются, что снижает интенсивность фотосинтеза.
Засухоустойчивость растений Засуха — это длительный бездождливый период, сопровождаемый снижением относительной влажности воздуха, влажности почвы и повышением температуры, когда не обеспечиваются нормальные потребности растений в воде. Засухоустойчивость — способность растений переносить длительные засушливые периоды, значительный водный дефицит, обезвоживание клеток, тканей и органов. При этом ущерб урожая зависит от продолжительности засухи и ее напряженности. Различают засуху почвенную и атмосферную.
Классификация ферментов
По типу катализируемых реакций ферменты подразделяются на 6 классов согласно иерархической классификации ферментов.
1: Оксидоредуктазы, катализирующие окисление или восстановление. Пример: каталаза, алкогольдегидрогеназа. 2: Трансферазы, катализирующие перенос химических групп с одной молекулы субстрата на другую. Среди трансфераз особо выделяют киназы, переносящие фосфатную группу, как правило, с молекулы АТФ. 3: Гидролазы, катализирующие гидролиз химических связей. Пример: эстеразы, пепсин, трипсин, амилаза, липопротеинлипаза. 4: Лиазы, катализирующие разрыв химических связей без гидролиза с образованием двойной связи в одном из продуктов. 5: Изомеразы, катализирующие структурные или геометрические изменения в молекуле субстрата. 6: Лигазы, катализирующие образование химических связей между субстратами за счёт гидролиза АТФ. Пример: ДНК-полимераза.
Клеточные мембраны Клеточная мембрана- отделяет содержимое любой клетки от внешней среды, обеспечивая её целостность; регулирует обмен между клеткой и средой; 7. ферментативная — мембранные белки нередко являются ферментами (белки, выполняющие роль катализаторов в живых организмах.) 35.Крахмал: физиологическая роль, биосинтез и распад в растении
Крахмал(С6Н10О5)n представляет собой природный полимер. Причем крахмал не индивидуальное вещество, а смесь двух полимеров состава – амилозы (10–20 %) и амилопектина (80–90 %), состоящих из остатков a-D-глюкозы. Амилоза- длинные не разветвленные цепи, сост из глюкопиранозы, соед между 1 и 4 углеродным атомом. Легко растворяется в теплой воде, растворы не устойчивы и могут образовывать остатки. С йодом окрашивается в синий цвет. Амилопектин-состоит из a-D глюкопиранозы. Связи которые имеются между 1, 4 и 1, 6 атомами углерода. Точки ветвления образуются через 25-30 остатков глюкозы. Крахмал, являясь одним из продуктов фотосинтеза, широко распространен в природе. Для растений он является запасом питательных веществ и содержится в основном в плодах, семенах и клубнях. Наиболее богато крахмалом зерно злаковых растений: риса (до 86 %), пшеницы (до 75 %), кукурузы (до 72 %), а также клубни картофеля (до 24 %). В пищевой промышленности крахмал используется для получения глюкозы, патоки, этанола, в текстильной — для обработки тканей, в бумажной — в качестве наполнителя.
БИОСИНТЕЗ: 1.Для образования амилазы необходимо: Распад крахмала происходит при участии двух процессов - гидролиза и фосфоролиза. Гидролитический распад крахмала осуществляется под действием четырех ферментов класса гидролиз α -амилаза, катализирует расщепление α (1-4)-связи, причем связи разрываются беспорядочно. Конечный продукт такого распада — мальтоза, глюкоза, декстрины. Под действием β -амилазы происходит расщепление α (1-4)-связей с образованием остатков мальтозы. Фермент глюкоамилазы катализирует последовательное отщепление остатков глюкозы от молекулы крахмала. Амилопектин-1, 6-глюкозидаза или R-фермент катализирует расщепление α (1-6)-связей в молекуле амилопектина, т. е. действует на точки ветвления. Фосфоролиз — это присоединение фосфорной кислоты по месту разрыва глюкозидной связи между остатками моносахаридов в цепи полисахарида, при этом происходит образование глюкозо-1-фосфата. Эта реакция катализируется ферментомаглюконфосфорилазой, относящимся к классу трансфераз. Крахмал в растении может подвергаться очень быстрому распаду, так как ферменты распада находятся во всех органах растения. 36.Лиазы, лигазы и их роль в растении.
ЛИАЗЫ, класс ферментов, катализирующих реакции, в результате которых происходит разрыв связи( С—С, С—О, С—N и др)сопровождающийся образованием двойных связей, а также обратные реакции присоединения по двойным связям. В названии фермента присутствуют слова " декарбоксилаза" и " альдолаза" или " лиаза" а для ферментов, катализирующих реакции отщепления воды от субстрата - " дегидратаза" (карбонат-дегидратаза, цитрат-дегидратаза, серин-дегидратаза и др.). В тех случаях, когда обнаружена только обратная реакция, или это направление в реакциях более существенно, в названии ферментов присутствует слово " синтаза" (малат-синтаза, 2-изопропилмалат-синтаза, цитрат-синтаза и др). ЛИГАЗЫ (латин. ligā re — сшивать, соединять) — фермент, катализирующий соединение двух молекул с образованием новой химической связи ( лигирование ). При этом обычно происходит отщепление (гидролиз) небольшой химической группы от одной из молекул.
37.Лист как основной фотосинтезирующий орган.
Лист растения - это основной орган растения, где проходит процесс фотосинтеза. Поскольку в основном лист покрыт малопроницаемой для газов кутикулой, то поступление СО2 в ткани идет через устьица, а в тканях - через сильно разветвленную сеть межклеточных воздухоносных каналов. К верхней стороне листа прилегает палисадная паренхима, клетки которой расположены перпендикулярно, плотно соприкасаются друг с другом и содержат много хлоропластов. Эта палисадная паренхима и является основной ассимиляционной тканью. К нижнему эпидермису прилегает губчатая паренхима с рыхло расположенными клетками и межклетниками. Кроме того, весь лист пронизан жилками, по которым идет перенос веды, минеральных ионов и ассимилятов. В палисадной паренхиме нет ни одной клетки, которая отстояла бы от ближайшей к ней жилки дальше, чем на несколько клеточных диаметров. Пройдя сквозь устьичный барьер, атмосферный СО2 растворяется в воде, гидратируется и превращается в угольную кислоту, а затем диссоциирует до бикарбонат-ионов (НСО3+), запас которых и служит резервом потенциального СО2 для использования в фотосинтезе. Поскольку основной тканью, поглощающей энергию солнца, является палисадная паренхима, содержащая максимальное количество хлоропластов, то, зная соотношение между площадью листа и площадью поверхности хлоропластов, можно приблизительно определить и поглощающую способность посевов. Так, на 1 га посева в среднем приходится 5 га листовой поверхности, то есть 1000 га поверхности хлоропластов, так как 1 см2листовой поверхности соответствует 200 см2 поверхности хлоропластов. При этом площадь поверхности межклетников, испаряющих воду составляет 50 га. В этом проявляется общебиологический закон - создание внутренних рабочих поверхностей при сравнительно малых наружных испаряющих площадях за счет затрат небольших количеств материала. В зависимости от условий обитания растений (засушливый или избыточно влажный климат, тропический климат с чрезмерной интенсивностью солнечного излучения) в строении листьев могут наблюдаться те или иные морфологические или биохимические особенности, однако общие принципы строения листа сохраняются. 38.Макроэргические соединения растительной клетки и их роль в жизни растений.
К макроэргическим соединениям относится определенный тип веществ, в ходе превращения которых происходит большее изменение свободной энергии, составляющее в стандартных условиях 30-60кДж/моль. В состав любого макроэргического соединения входит группировка атомов, присоединенная сильно полярной связью, которую обозначают специальном символом ” ~” и называют макроэргической связью. В зависимости от макроэргической связи различают 3 группировки макроэргических соединений: 1.Фосфаты –содержат остатки ортофосфорной кислоты, соединенные макроэргической связью с остатками нуклеотидов (нуклеозидполифосфаты), карбоновых кислот(ацилфосфаты), аминными (амидинфосфаты) и енольными (енолфосфаты) производными. · АТФ(аденозинтрифосфат), ГТФ(гуанозинтрифосфат), УТФ(уридинтрифосфат), ЦТФ(цитидинтрифосфат). · Ацилфосфатом является 1, 3-дифосфоглицериновая кислота · Енолфосфатом является фосфоенолпировиноградная кислота · Амидинфосфатом является аргининфосфат
2.Тиоэфиры-ацильные производные CoА.Важнейшим является ацетил CoA. Другие макроэргические тиоэфиры отличаются от ацетила CoA длинной цепи углеводородного радикала, характерной для конкретной карбоновой кислоты(пропионилкофермент А, малонилкофермент А и др.) 3.Имидазолы. 39 Митохондрии: строение и функции
Митохондрия – двумембранный полуавтономный органоид, осуществляющий синтез АТФ.Форма митохондрий разнообразна, они могут быть палочковидными, нитевидными или шаровидными. Стенки митохондрий образованы двумя мембранами: внешней и внутренней. Внешняя мембрана - гладкая, а внутренняя образует многочисленные складки - кристы. Во внутренней мембране встроены многочисленные ферментные комплексы, которые осуществляют синтез АТФ. Митохондрии называют полуавтономными органоидами. Это означает, что они зависят от клетки, но в то же время сохраняют некоторую самостоятельность. Так, например, митохондрии сами синтезируют собственные белки, в том числе и ферменты своих ферментных комплексов. Кроме того, митохондрии могут размножаться путем деления независимо от деления клетки. Внутреннее пространство митохондрий заполнено бесструктурным однородным веществом (матриксом). В матриксе располагаются кольцевые молекулы ДНК, РНК и мелкие рибосомы (как у прокариот). В ДНК митохондрий записана информация о строении митохондриальных белков. РНК и рибосомы осуществляют их синтез. Рибосомы митохондрий мелкие, по строению они очень похожи на рибосомы бактерий.. Складчатость внутренней мембраны имеет большое значение. На складчатой поверхности может расположиться больше ферментных комплексов, чем на гладкой поверхности. Количество складок в митохондрии может изменяться в зависимости от потребности клеток в энергии.Если клетка нуждается в энергии, то число крист увеличивается. Соответственно увеличивается и число ферментных комплексов, расположенных на кристах. В результате будет образовано большее количество АТФ. Кроме того, в клетке может возрастать общее количество митохондрий. Если клетка не нуждается в большом количестве энергии, то количество митохондрий в клетке снижается и уменьшается количество крист внутри митохондрий. Основная функция — окисление органических соединений и использование освобождающейся при их распаде энергии в синтеземолекул АТФ, который происходит за счёт движения электрона по электронно-транспортной цепи белков внутренней мембраны. 40.Моносахориды, их фосфорные эфиры, биосинтез и превращение в растениях. Значение моносахоридов.
Моносахариды(триозы, тетрозы, пентозы, гексозы, гектозы) - бесцветные кристаллические вещества, хорошо рстворимые в воде, плохо - в спирте, нерастворимые в эфире. Моносахариды - основной источник энергии в организме человека. Моносахариды представляют собой соединения со смешанными функциями из-за наличия в их структуре различных реакционноспособных групп. Моносахариды нелетучи, легко растворяются в воде и других полярных растворителях, что связано с наличием в их структуре большого количества полярных гидроксильных групп. Большинство из них легко кристаллизируются, устойчивы при воздействии разведенных кислот. Самый важный моносахарид - глюкоза. Название произошло от греческого - glykys - сладкий. Химическая формула - C6H12O6. Молекулы глюкозы выполняют роль биологического топлива в одном из важнейших энергегетических процессов в организме - в процессе гликолиза. В пентозном цикле глюкоза окисляется до СО2 и воды, генерируя энергию для некоторых реакций. В природе встречается D - глюкоза. Глюкоза очень легко окисляется оксидами и гидроксидами тяжелых металлов. Полное окисление глюкозы идет по уравнению: C6H12O6 + 6O2 = 6CO2 + 6 H2O + 686 ккал. - урановые кислоты – глюкуроновая, галактуроновая, аскорбиновая кислоты. Очень часто они входят в состав протеогликанов; - аминосахара – глюкозамин, галактозамин. Ряд антибиотиков (эритромицин, карбомицин) содержат в своём составе аминосахара; - сиаловые кислоты. Входят в состав протеогликанов и гликолипидов; - гликозиды – примером служат сердечные гликозиды, антибиотик стрептомицин. Биосинтез моносахаридов из углекислого газа и воды происходит в растениях с участием активированных производных моносахаридов — нуклеозиддифосфатсахаров — происходит, как правило, биосинтез сложных углеводов. Распад моносахаридов в организме (например, спиртовое брожение, гликолиз) сопровождается выделением энергии.
Морозоустойчивость растений
Морозоустойчивость растений, способность растений переносить температуру ниже 0˚ С и низкие отрицательные температуры. Морозоустойчивые растения способны предотвращать или уменьшать действие низких отрицательных температур. Растения переносят условия зимы в различные периоды онтогенеза. У однолетних культур зимуют семена (яровые растения), раскустившиеся растения (озимые), у двулетних и многолетних — клубни, корнеплоды, луковицы, корневища, взрослые растения. Способность озимых, многолетних травянистых и древесных плодовых культур перезимовывать обусловливается их достаточно высокой морозоустойчивостью. Ткани этих растений могут замерзать, однако растения не погибают.
Фазы закаливания: Первая фаза закаливания проходит на свету и при низких положительных температурах в ночное время (днем около 10 °С, ночью около 2 °С), останавливающих рост, и умеренной влажности почвы. Озимые злаки проходят первую фазу на свету при среднесуточной температуре 0, 5-2 °С за 6-9 дней, древесные — за 30 дней. В эту фазу продолжается дальнейшее замедление и даже происходит полная остановка ростовых процессов. Вторая фаза закаливания не требует света и начинается сразу же после первой фазы при температуре немного ниже 0 °С. Для травянистых растений она может протекать и под снегом. Длится она около двух недель при постепенном снижении температуры до -10…-20 °С и ниже со скоростью 2-3 °С в сутки, что приводит к частичной потере воды клетками, освобождению клеток тканей от избыточного содержания воды или витрификации (переходу воды в стеклообразное состояние). Явление витрификации воды в растительных клетках наступает при резком охлаждении (ниже -20 °С). Стеклообразная растительная ткань долго сохраняет свою жизнеспособность. 42.Настии и их роль в растении
Настии – движения органов растения, вызываемые раздражителями, действующими равномерно на все растения, например изменениями температуры, влажности, освещенности и т. п. Классификация настий: Настии играют большую роль в жизни растений, это один из признаков приспособленности растений к среде обитания, к активной реакции на воздействие различных ее факторов. Показатели транспирации Транспирация – это физиологический процесс испарения воды растением. Транспирация необходима: 1. транспирация спасает растение от перегрева, который ему грозит на прямом солнечном свете. Температура транспирирующего листа на 5-7 градусов ниже температуры окружающего воздуха; 3. транспирация создает непрерывный ток воды из корневой системы к листьям и связывает все органы растения в единое целое; 4. с транспирационным током передвигаются растворимые минеральные и частично органические питательные вещества, при этом интенсивнее транспирация, тем быстрее идет процесс.
Значение транспирации: - является верхним двигателем водного тока; - передвижение воды по растению; - связано с поступлением CO2; - влияет на метаболизм в растении; - влияет на температуру растения.
Интенсивность транспирации – величина, показывающая, сколько граммов воды испарилось с единицы площади за единицу времени (меняется от 1г до 250 г). Транспирационный эффект – число г воды при образовании 1 г сухого вещества (от 125 г до 1000 г). Зависит от вида растений, ярусности листьев, условий внешней среды. Продуктивность транспирации – показывает, сколько г сухого вещества образуется при расходе 1 кг воды (от 1 до 8 г). Относительная транспирация – отношение интенсивности транспирации к интенсивности испарения со свободной поверхности (от 0, 1 г до 1 г). Регулирование устьичной транспирации – осуществляется открытием или закрытием устьиц. Их движение обусловлено различными факторами. Как мы уже отмечали, основным, обусловливающим движением устьиц является содержание воды в замыкающих клетках (изменение тургора). Различают гидропассивное и гидроактивное открытие и закрытие устьиц. Гидропассивная реакция – это закрытие устьичных щелей, вызванное тем, что окружающие паренхимные клетки, переполненные водой, механически сдавливают замыкающие клетки. В результате сдавливания устьица не могут открыться. Гидропассивное движение обычно наблюдается после сильных поливов и может служить причиной торможения процесса фотосинтеза, а также скажется на тех процессах, которые связаны с током воды по растению. Гидроактивная реакция открывания и закрывания – это движение замыкающих клеток, вызванное применением содержания воды. Это связано с изменением концентрации осмотически активных веществ в процессе фотосинтеза, в замыкающих клетках.
Факторы, влияющие на транспирацию: 1. С повышением температуры транспирация возрастает. 2. На свету зеленые листья поглощают определенные участки спектра, повышается температура листа и, следовательно, усиливается процесс транспирации. Действие света на транспирацию усиливается тем больше, чем выше содержание хлорофилла. На свету увеличивается проницаемость цитоплазмы. 4. Интенсивность транспирации зависит и от ряда внутренних факторов, и прежде всего от содержания воды в листьях. Всякое уменьшение содержания воды в листьях уменьшает транспирацию. 5. Транспирация зависит и от концентрации клеточного сока. Чем концентрированнее клеточный сок, тем слабее транспирация. Интенсивность транспирации зависит от эластичности клеточных стенок. 7. На процесс транспирации влияет смена дня и ночи. В ночной период суток транспирация резко сокращается из-за снижения температуры, повышения влажности воздуха, отсутствия света. 9. Транспирация зависит от величины листовой поверхности, чем она (листовая поверхность) больше, тем сильнее процесс транспирации.
Минеральные вещества. 1) К+, Cu2+, Fe2+, Fe3+ 2) Cl-, SO42-, PO43- Лучше поступают катионы (т.к. заряд клеточной стенки к внешней среде отрицательный). Чем меньше заряд, тем легче проходят. Органические вещества. 1) неполярные группировки: СН2, СН3 2) полярные группировки: ОН, СООН Поступление органических веществ не зависит от молекулярной массы, а определяется составом. Чем больше неполярных группировок, тем легче поступают.
Проницаемость цитоплазмы, поступление молекул и ионов в клетку — это активный процесс обмена веществ. Под влиянием растворов солей одновалентных металлов (натрий и калий) проницаемость цитоплазмы значительно увеличивается. А под влиянием двухвалентных ионов металлов (кальций и, реже, магний), соответственно, уменьшается.
Таким образом, поддержание проницаемости цитоплазмы на определенном уровне связано с сохранением баланса между ионами содержащимися в окружающей клетку среде и в самой цитоплазме. Их соотношение определяет вязкость — одна из важнейших характеристик цитоплазмы. Причем, концентрация отдельных ионов в клетке и в среде не бывает одинаковой. Поступают ионы в клетку путем пассивного не метаболического поглощения, в результате диффузии, по градиенту концентрации и, соответственно, кривой насыщения. И путем метаболической активной абсорбции. Плазматические мембраны хорошо пропускают воду и газы и малопроницаемы для ионов металлов, которые поглощаются и против градиента концентрации. Аммонификация. Мочевая кислота, выделяемая птицами и рептилиями, также быстро минерализуется особыми группами микроорганизмов с образованием NH3 и СО2. С другой стороны, азот, включённый в состав живых существ, после их гибели подвергается аммонификации (разложение содержащих азот сложных соединений с выделением аммиака и ионов аммония (NH4+)) и нитрификации.
Денитрификация. Продукты нитрификации — NO3- и (NO2-) в дальнейшем подвергаются денитрификации. Диссимиляционное восстановление нитрата является процессом анаэробного дыхания, то есть использования нитратов и продуктов их частичного восстановления вместо кислорода для окисления веществ (у разных микроорганизмов как органических, так и минеральных) в ходе метаболизма с выделением энергии.
Ассимиляция. Усваиваемые соединения азота могут накапливаться в почве в неорганической форме (нитрат) или могут быть включены в живой организм как органический азот. Ассимиляция и минерализация определяет поглощение соединений азота из почвы, объединение их в биомолекулы растений и конверсию в неорганический азот после отмирания растений, соответственно. Ассимиляция - переход неорганического азота (типа нитрата) в органическую форму азота как, например, аминокислоты. Нитрат переходит с помощью ферментов сначала в нитрит (редуктаза нитрата), затем в аммиак (редуктаза нитрита). Аммиак входит в состав аминокислот.
Образование лизосом По морфологии выделяют 4 типа лизосом: 1. Первичные лизосомы 2. Вторичные лизосомы 3. Аутофагосомы 4. Остаточные тельца
Первичные лизосомы представляют собой мелкие мембранные пузырьки, заполненные бесструктурным веществом, содержащим набор гидролаз. Маркерным ферментом для лизосом является кислая фосфотаза. Первичные лизосомы настолько мелкие, что их очень трудно отличить от мелких вакуолей на периферии зоны аппарата Гольджи. В дальнейшем первичные лизосомы сливаются с фагоцитарными или пиноцитарными вакуолями и образуют вторичные лизосомы или внутриклеточная пищеварительная вакуоль. При этом содержимое первичной лизосомы сливается с содержимым фагоцитарной или пиноцитарной вакуолей, а гидролазы первичной лизосомы получают доступ к субстратам, которые они начинают расщеплять. Лизосомы могут сливаться друг с другом и таким путем увеличиваться в объеме, при этом усложняется их внутренняя структура. Судьба веществ, попавшивших в лизосомы, заключается в их расщеплении гидролазами до мономеров, мономеры транспортируются через мембрану лизосомы в гиалоплазму, где включаются в различные обменные процессы. Расщепление и переваривание может идти не до конца. В этом случае в полости лизосом накапливаются непереваренные продукты, и вторичные лизосомы переходят в остаточные тельца. Остаточные тельца содержат меньше гидролитических ферментов, в них происходит уплотнение содержимого и его переотработка. Часто в остаточных тельцах наблюдается вторичная структуризация непереваренных липидов, которые образуют сложные слоистые структуры. Происходит отложение пигментных веществ. Аутофагосомы встречаются в клетках простейших. Они относятся к вторичным лизосомам. Но в своем состояние содержат фрагменты цитоплазматических структур (остатки митохондрий, пластид, ЭПР, остатки рибосом, так же могут содержать гранулы гликогена). Процесс образования не ясен, но предполагают, что первичные лизосомы выстраиваются вокруг клеточной органеллы, сливаются друг с другом и отделяют органеллу от соседних участков цитоплазмы. Предполагают, что аутофагоцитоз связан с уничтожением сложных клеточных компонентов. В нормальных условиях число аутофагосом возрастает при метаболических стрессах. При различных повреждениях клеток аутофагоцитозу могут подвергаться целые зоны клеток. Лизосомы присутствуют в самых разных клетках. Некоторые специализированны клетки, например лейкоциты, содержат их в особенно большом количестве. Интересно, что отдельные виды растений, в клетках которых лизосомы не обнаружены, содержат гидролитические ферменты в клеточных вакуолях, которые поэтому могут выполнять ту же функцию, что и лизосомы. Функция лизосом, по-видимому, лежит в основе таких процессов, автолиз и некроз тканей, когда ферменты освобождаются из этих органелл в результате случайных или «запрограммированных» процессов. Естественной функцией лизосом является поставка гидролитических ферментов как для внутриклеточного, так и, возможно, для внеклеточного использования; после слияния мембран содержимое лизосом может смешиваться с содержимым фагоцитозных пузырьков, так что процессы гидролиза протекают в пространстве, обособленном от всех областей цитоплазмы, в которых находятся уязвимые для гидролиза внутриклеточные компоненты. Показано, что лизосомные ферменты могут освобождаться и во внеклеточное пространство. Продукты гидролиза могут проникать из органеллы в цитоплазму или выводиться из клетки наружу. Комплекс Гольджи представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям, и связанную с ними систему пузырьков Гольджи. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько, соединённых трубками, стопок. В Комплексе Гольджи выделяют 3 отдела цистерн, окружённых мембранными пузырьками: 1.Цис-отдел (ближний к ядру); 2.Медиальный отдел; 3.Транс-отдел (самый отдалённый от ядра). Эти отделы различаются между собой набором ферментов. В цис-отделе первую цистерну называют " цистерной спасения", так как с её помощью рецепторы, поступающие из промежуточной эндоплазматической сети, возвращаются обратно. Фермент цис-отдела: фосфогликозидаза (присоединяет фосфат к углеводу - маннозе). В медиальном отделе находится 2 фермента: манназидаза (отщепляет манназу) и N-ацетилглюкозаминтрансфераза (присоединяет определенные углеводы - гликозамины). В транс-отделе ферменты: пептидаза (осуществляет протеолиз) и трансфераза (осуществляет переброс химических групп). Функции 1.Разделение белков на 3 потока: Популярное:
|
Последнее изменение этой страницы: 2016-04-11; Просмотров: 2678; Нарушение авторского права страницы