|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Роль печени в обмене углеводов.Стр 1 из 5Следующая ⇒
Биохимия печени Функции печени. Печень является органом, занимающим уникальное место в обмене веществ. В каждой печёночной клетке содержится несколько тысяч ферментов, катализирующих реакции многочисленных метаболических путей. Поэтому печень выполняет в организме целый ряд метаболических функций. Важнейшими из них являются: · биосинтез веществ, которые функционируют или используются в других органах. К этим веществам относятся белки плазмы крови, глюкоза, липиды, кетоновые тела и многие другие соединения; · биосинтез конечного продукта азотистого обмена в организме - мочевины; · участие в процессах пищеварения - синтез желчных кислот, образование и экскреция желчи; · биотрансформация (модификация и конъюгация) эндогенных метаболитов, лекарственных препаратов и ядов; · выделение некоторых продуктов метаболизма (желчные пигменты, избыток холестерола, продукты обезвреживания).
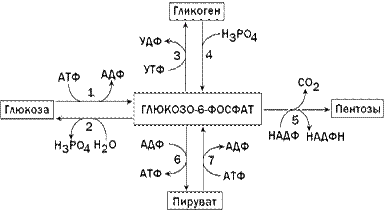
Роль печени в обмене углеводов. Основная роль печени в обмене углеводов заключается в поддержании постоянного уровня глюкозы в крови. Это осуществляется путём регуляции соотношения процессов образования и утилизации глюкозы в печени. В клетках печени содержится фермент глюкокиназа, катализирующий реакцию фосфорилирования глюкозы с образованием глюкозо-6-фосфата. Глюкозо-6-фосфат является ключевым метаболитом углеводного обмена; основные пути его превращения представлены на рисунке 1.
31.2.1. Пути утилизации глюкозы. После приёма пищи большое количество глюкозы поступает в печень по воротной вене. Эта глюкоза используется прежде всего для синтеза гликогена (схема реакций приводится на рисунке 2). Содержание гликогена в печени здоровых людей обычно составляет от 2 до 8% массы этого органа. Гликолиз и пентозофосфатный путь окисления глюкозы в печени служат в первую очередь поставщиками метаболитов-предшественников для биосинтеза аминокислот, жирных кислот, глицерола и нуклеотидов. В меньшей степени окислительные пути превращения глюкозы в печени являются источниками энергии для обеспечения биосинтетических процессов.
Рисунок 1. Главные пути превращения глюкозо-6-фосфата в печени. Цифрами обозначены: 1 - фосфорилирование глюкозы; 2 - гидролиз глюкозо-6-фосфата; 3 - синтез гликогена; 4 - мобилизация гликогена; 5 - пентозофосфатный путь; 6 - гликолиз; 7 - глюконеогенез.
Рисунок 2. Схема реакций синтеза гликогена в печени.
Рисунок 3. Схема реакций мобилизации гликогена в печени.
31.2.2. Пути образования глюкозы. В некоторых условиях (при голодании низкоуглеводной диете, длительной физической нагрузке) потребность организма в углеводах превышает то количество, которое всасывается из желудочно-кишечного тракта. В таком случае образование глюкозы осуществляется с помощью глюкозо-6-фосфатазы, катализирующей гидролиз глюкозо-6-фосфата в клетках печени. Непосредственным источником глюкозо-6-фосфата служит гликоген. Схема мобилизации гликогена представлена на рисунке 3. Мобилизация гликогена обеспечивает потребности организма человека в глюкозе на протяжении первых 12 - 24 часов голодания. В более поздние сроки основным источником глюкозы становится глюконеогенез - биосинтез из неуглеводных источников. Основными субстратами для глюконеогенеза служат лактат, глицерол и аминокислоты (за исключением лейцина). Эти соединения сначала превращаются в пируват или оксалоацетат - ключевые метаболиты глюконеогенеза. Глюконеогенез - процесс, обратный гликолизу. При этом барьеры, создаваемые необратимыми реакциями гликолиза, преодолеваются при помощи специальных ферментов, катализирующих обходные реакции (см. рисунок 4). Из других путей обмена углеводов в печени следует отметить превращение в глюкозу других пищевых моносахаридов - фруктозы и галактозы.
Рисунок 4. Гликолиз и глюконеогенез в печени. Ферменты, катализирующие необратимые реакции гликолиза: 1 - глюкокиназа; 2 - фосфофруктокиназа; 3 - пируваткиназа. Ферменты, катализирующие обходные реакции глюконеогенеза: 4 -пируваткарбоксилаза; 5 - фосфоенолпируваткарбоксикиназа; 6 -фруктозо-1, 6-дифосфатаза; 7 - глюкозо-6-фосфатаза.
Рисунок 5. Образование и использование ацетил-КоА в печени.
31.3.1. Метаболизм жирных кислот в печени. Пищевые жиры в виде хиломикронов поступают в печень через систему печёночной артерии. Под действием липопротеинлипазы, находящейся в эндотелии капилляров, они расщепляются до жирных кислот и глицерола. Жирные кислоты, проникающие в гепатоциты, могут подвергаться окислению, модификации (укорочению или удлинению углеродной цепи, образованию двойных связей) и использоваться для синтеза эндогенных триацилглицеролов и фосфолипидов. 31.3.2. Синтез кетоновых тел. При β -окислении жирных кислот в митохондриях печени образуется ацетил-КоА, подвергающийся дальнейшему окислению в цикле Кребса. Если в клетках печени имеется дефицит оксалоацетата (например, при голодании, сахарном диабете), то происходит конденсация ацетильных групп с образованием кетоновых тел (ацетоацетат, β -гидроксибутират, ацетон). Эти вещества могут служить энергетическими субстратами в других тканях организма (скелетные мышцы, миокард, почки, при длительном голодании - головной мозг). Печень не утилизирует кетоновые тела. При избытке кетоновых тел в крови развивается метаболический ацидоз. Схема образования кетоновых тел - на рисунке 6.
Рисунок 6. Синтез кетоновых тел в митохондриях печени. 31.3.3. Образование и пути использования фосфатидной кислоты. Общим предшественником триацилглицеролов и фосфолипидов в печени является фосфатидная кислота. Она синтезируется из глицерол-3-фосфата и двух ацил-КоА - активных форм жирных кислот (рисунок 7). Глицерол-3-фосфат может образоваться либо из диоксиацетонфосфата (метаболит гликолиза), либо из свободного глицерола (продукт липолиза).
Рисунок 7. Образование фосфатидной кислоты (схема).
Для синтеза фосфолипидов (фосфатидилхолина) из фосфатидной кислоты необходимо поступление с пищей достаточного количества липотропных факторов (веществ, препятствующих развитию жировой дистрофии печени). К этим факторам относятся холин, метионин, витамин В12, фолиевая кислота и некоторые другие вещества. Фосфолипиды включаются в состав липопротеиновых комплексов и принимают участие в транспорте липидов, синтезированных в гепатоцитах, в другие ткани и органы. Недостаток липотропных факторов (при злоупотреблении жирной пищей, хроническом алкоголизме, сахарном диабете) способствует тому, что фосфатидная кислота используется для синтеза триацилглицеролов (нерастворимых в воде). Нарушение образования липопротеинов приводит к тому, что избыток ТАГ накапливается в клетках печени (жировая дистрофия) и функция этого органа нарушается. Пути использования фосфатидной кислоты в гепатоцитах и роль липотропных факторов показаны на рисунке 8.
Рисунок 8. Использование фосфатидной кислоты для синтеза триацилглицеролов и фосфолипидов. Липотропные факторы обозначены знаком *.
31.3.4. Образование холестерола. Печень является основным местом синтеза эндогенного холестерола. Это соединение необходимо для построения клеточных мембран, является предшественником желчных кислот, стероидных гормонов, витамина Д3. Первые две реакции синтеза холестерола напоминают синтез кетоновых тел, но протекают в цитоплазме гепатоцита. Ключевой фермент синтеза холестерола - β -гидрокси-β -метилглутарил-КоА-редуктаза (ГМГ-КоА-редуктаза) ингибируется избытком холестерола и желчными кислотами по принципу отрицательной обратной связи (рисунок 9).
Рисунок 9. Синтез холестерола в печени и его регуляция.
31.3.5. Образование липопротеинов. Липопротеины - белково-липидные комплексы, в состав которых входят фосфолипиды, триацилглицеролы, холестерол и его эфиры, а также белки (апопротеины). Липопротеины транспортируют нерастворимые в воде липиды к тканям. В гепатоцитах образуются два класса липопротеинов - липопротеины высокой плотности (ЛПВП) и липопротеины очень низкой плотности (ЛПОНП).
Углеводный обмен В гепатоцитах активно протекают процессы углеводного обмена. Благодаря синтезу и распаду гликогена печень поддерживает концентрацию глюкозы в крови. Активный синтез гликогена происходит после приема пищи, когда концентрация глюкозы в крови воротной вены достигает 20 ммоль/л. Запасы гликогена в печени составляют от 30 до 100 г. При кратковременном голодании происходит гликогенолиз, в случае длительного голодания основным источником глюкозы крови является глюконеогенез из аминокислот и глицерина. Печень осуществляет взаимопревращение сахаров, т.е. превращение гексоз (фруктозы, галактозы) в глюкозу. Активные реакции пентозофосфатного пути обеспечивают наработку НАДФН, необходимого для микросомального окисления и синтеза жирных кислот и холестерола из глюкозы. Липидный обмен Если во время приема пищи в печень поступает избыток глюкозы, который не используется для синтеза гликогена и других синтезов, то она превращается в липиды – холестерол и триацилглицеролы. Поскольку запасать ТАГ печень не может, то их удаление происходит при помощи липопротеинов очень низкой плотности ( ЛПОНП ). Холестерол используется, в первую очередь, для синтеза желчных кислот, также он включается в состав липопротеинов низкой плотности ( ЛПНП ) и ЛПОНП. При определенных условиях – голодание, длительная мышечная нагрузка, сахарный диабет I типа, богатая жирами диета – в печени активируется синтез кетоновых тел, используемых большинством тканей как альтернативный источник энергии. Белковый обмен Больше половины синтезируемого за сутки в организме белка приходится на печень. Скорость обновления всех белков печени составляет 7 суток, тогда как в других органах эта величина соответствует 17 суткам и более. К ним относятся не только белки собственно гепатоцитов, но и идущие на " экспорт" – альбумины, многие глобулины, ферменты крови, а также фибриноген и факторы свертывания крови. Аминокислоты подвергаются катаболическим реакциям с трансаминированием и дезаминированием, декарбоксилированию с образованием биогенных аминов. Происходят реакции синтеза холина и креатина благодаря переносу метильной группы от аденозилметионина. В печени идет утилизация избыточного азота и включение его в состав мочевины. Реакции синтеза мочевины теснейшим образом связаны с циклом трикарбоновых кислот. Тесное взаимодействие синтеза мочевины и ЦТК Пигментный обмен Участие печени в пигментном обмене заключается в превращении гидрофобного билирубина в гидрофильную форму и секреция его в желчь. Пигментный обмен, в свою очередь, играет важную роль в обмене железа в организме – в гепатоцитах находится железосодержащий белок ферритин. Оценка реакций 1-й фазы Оценку микросомального окисления можно проводить следующими способами: · определение активности микросомальных ферментов после биопсии, · по фармакокинетике препаратов, · с помощью метаболических маркеров ( антипириновая проба ). 5.2. Реакции конъюгации ксенобиотиков. К реакциям конъюгации относятся глюкуронидная, сульфатная, ацетильная, метильная и пептидная конъюгация.
Ксенобиотики (чужеродные соединения) - природные или синтетические вещества, не используемые в организме в качестве источников энергии или структурных компонентов тканей. К этой категории веществ могут быть отнесены многие лекарственные средства, а также соединения, применяемые для защиты растений, инсектициды, отходы промышленных предприятий, пищевые добавки, красящие вещества, вкусовые средства, консерванты, косметические составы.
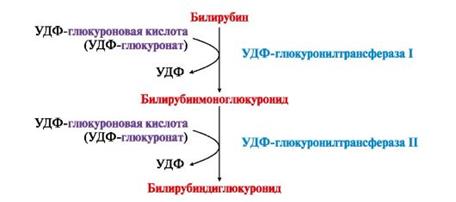
Глюкуронидная конъюгация. Реакцию катализирует глюкуронилтрансфераза, коферментом является активная форма глюкуроновой кислоты - уридин-дифосфоглюкуроновая кислота (УДФ-глюкуроновая кислота). В реакцию вступают спирты, фенолы, карбоновые кислоты, тиолы и амины. Из эндогенных субстратов можно отметить билирубин, стероидные гормоны, витамин D. Примером реакции может служить образование фенилглюкуронида:
Сульфатная конъюгация. Реакцию катализирует сульфотрансфераза. В качестве активной формы сульфата выступает З-фосфоаденозин-5-фосфосульфат (ФАФС). Субстратами чаще всего служат спирты и фенолы, реже - аминосоединения. В качестве примера реакции можно привести конъюгацию индоксила, который образуется в результате гидроксилирования индола(см. 33.5.1., реакции гидроксилирования ароматических соединений):
Продукт этой реакции в виде калиевой соли (животный индикан) выводится почками. Определение содержания индикана в моче может быть использовано для оценки интенсивности процессов гниения белков в кишечнике.
Ацетильная конъюгация. Ацетилированием называют присоединение к молекуле ксенобиотика или его метаболита остатка уксусной кислоты. Ацетилированию подвергаются вещества, содержащие свободную аминогруппу (алифатические и ароматические амины, аминокислоты, гидразины, гидразиды). Из эндогенных субстратов можно упомянуть аминосахара (глюкозамин, галактозамин) и биогенные амины. Катализируют реакции ацетилирования ферменты ацетилтрансферазы, донором ацетильной группы является ацетил-КоА. Пример реакции - ацетилирование изониазида (изоникотиноилгидразида):
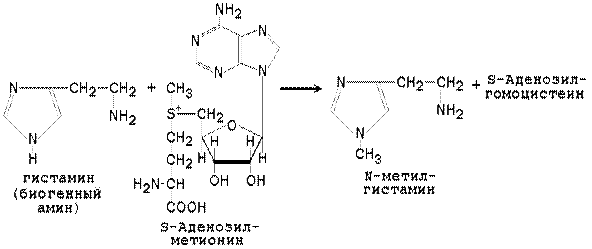
Метильная конъюгация (метилирование). Реакции метилирования (присоединения метильной группы) катализируют ферменты метилтрансферазы или трансметилазы. Донором метильной группы является активная форма аминокислоты метионина - S-аденозилметионин. Метилирование характерно для некоторых эндогенных субстратов (гуанидинацетат, норадреналин, фосфатидилэтаноламин). Субстратами для метилтрансфераз служат фенолы, тиолы и амины. Пример реакции - метилирование гистамина:
Метилирование ксенобиотиков по сравнению с другими реакциями конъюгации имеет одну особенность. В результате присоединения метильной группы продукт реакции не становится более гидрофильным. Тем не менее метильная конъюгация выполняет важную роль, так как в результате метилирования устраняются чрезвычайно реакционноспособные SH- и NН-группы. Пептидная конъюгация - взаимодействие ксенобиотиков или их метаболитов с аминокислотами (глицин, глутамин, таурин и др.) при помощи пептидных (амидных) связей. Особенность этой разновидности конъюгации заключается в том, что ксенобиотик вступает в реакцию в активной форме (в других типах конъюгации активируется биомолекула). Пептидная конъюгация характерна для соединений, содержащих карбоксильные группы. Примером может служить конъюгация бензойной кислоты с глицином, в результате чего образуется гиппуровая кислота:
Эта реакция лежит в основе пробы Квика, используемой для оценки обезвреживающей функции печени.
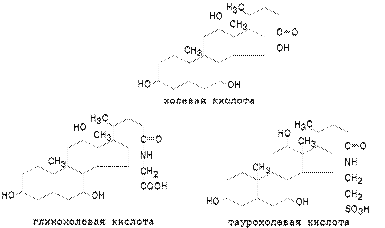
В реакции конъюгации с глицином (H2N-CH2-COOH) и таурином (H2N-CH2-CH2-SO3H) вступают также желчные кислоты (например, холевая), образуя " парные соединения" или конъюгаты.
Обмен билирубина
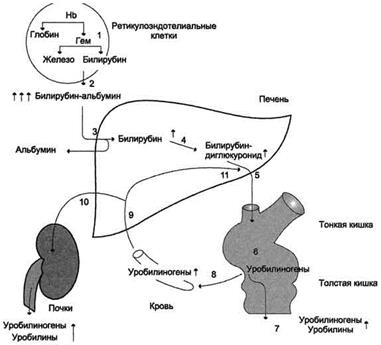
Гемолитическая желтуха Гемолитическая или надпеченочная желтуха – ускоренное образование билирубина в результате усиления внутрисосудистого гемолиза любого происхождения (сепсис, лучевое поражение, несовместимость крови, дефект глюкозо-6-фосфатдегидрогеназы). У новорожденных гемолитическая желтуха может развиться как симптом гемолитической болезни новорожденного. В данном случае гипербилирубинемия развивается за счет фракции непрямого билирубина. Гепатоциты усиленно переводят непрямой билирубин в связанную форму, секретируют его в желчь, в результате в кале увеличивается содержание стеркобилина, интенсивно его окрашивая. В моче возрастает содержание уробилина, билирубин отсутствует.
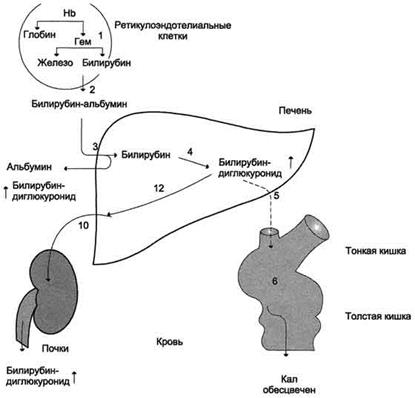
Рисунок 1. Билирубин-уробилиногеновый цикл при гемолитической желтухе. 1 - катаболизм Нb идёт с повышенной скоростью; 2 - в крови примерно в 10 раз повышена концентрация непрямого билирубина; 3 - альбумин высвобождается из комплекса билирубин-альбумин; 4 - активность реакции глюкуронирования возрастает, но она ниже, чем скорость образования билирубина; 5 - секреция билирубина в жёлчь повышена; 6, 7, 10 - повышенное содержание уробилиногенов в кале и моче придаёт им более интенсивную окраску; уробилиноген всасывается из кишечника в кровь (8) и снова попадает в печень по воротной вене (9). Механическая желтуха Механическая или подпеченочная желтуха развивается вследствие снижения оттока желчи при непроходимости желчного протока (закупорка желчного протока опухолями, желчными камнями). При этом происходит растяжение желчных капилляров, увеличивается проницаемость их стенок и не имеющий оттока в кишечник прямой билирубин поступает в кровь, развивается гипербилирубинемия. В моче резко увеличен уровень билирубина (цвет темного пива) и снижено количество уробилина, практически отсутствует стеркобилин кала (серовато-белое окрашивание).
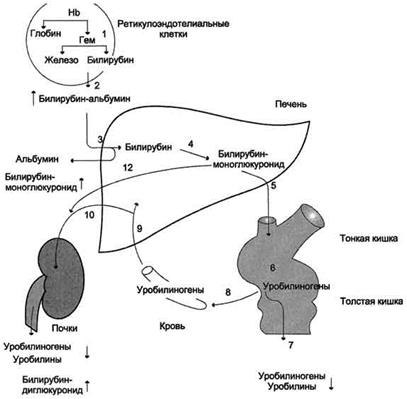
Рисунок 2. Нарушение билирубин-уробилиногенового цикла при обтурационной желтухе. Вследствие закупорки жёлчного пузыря билирубинглюкуронид не секретируется в жёлчь (5); отсутствие билирубина в кишечнике приводит к обесцвечиванию кала (6); растворимый билирубинглюкуронид выделяется почками с мочой (10). Уробилинов в моче нет; образующийся в печени билирубинглюкуронид поступает в кровь (12), вследствие этого возрастает содержание прямого билирубина. Паренхиматозная желтуха Паренхиматозная ( печеночно-клеточная ) желтуха – причиной может быть нарушение на всех трех стадиях превращения билирубина в печени: извлечение билирубина из крови печеночными клетками, его конъюгирование и секреция в желчь. Наблюдается при различных формах гепатитов (вирусные, токсические) и иных поражениях печени. Гипербилирубинемия развивается за счет обеих фракций. Их концентрация возрастает из-за одновременного нарушения секреции в желчь и увеличения проницаемости мембран клеток печени. В моче определяется билирубин (цвет крепкого черного чая), умеренно увеличена концентрация уробилина, уровень стеркобилина кала в норме или снижен. У младенцев вариантами паренхиматозной желтухи являются желтухи новорожденныхи недоношенных: физиологическая желтуха, желтуха, вызываемая молоком матери и т.п.. Наследственные желтухи печеночного происхождения представляют собой синдромы Жильбера-Мейленграхта, Дубина-Джонсона, Криглера-Найяра.
Рисунок 3. Нарушение билирубин-уробилиногенового цикла при печёночно-клеточной желтухе. В печени снижена скорость реакции глюкуронирования билирубина (4), поэтому в крови повышается концентрация непрямого билирубина; вследствие нарушения паренхимы печени часть образованного в печени билирубинглюкуронида попадает в кровь (12) и далее с мочой (10) удаляется из организма. В моче больных присутствуют уробилины и билирубинглюкурониды. Биохимия печени Функции печени. Печень является органом, занимающим уникальное место в обмене веществ. В каждой печёночной клетке содержится несколько тысяч ферментов, катализирующих реакции многочисленных метаболических путей. Поэтому печень выполняет в организме целый ряд метаболических функций. Важнейшими из них являются: · биосинтез веществ, которые функционируют или используются в других органах. К этим веществам относятся белки плазмы крови, глюкоза, липиды, кетоновые тела и многие другие соединения; · биосинтез конечного продукта азотистого обмена в организме - мочевины; · участие в процессах пищеварения - синтез желчных кислот, образование и экскреция желчи; · биотрансформация (модификация и конъюгация) эндогенных метаболитов, лекарственных препаратов и ядов; · выделение некоторых продуктов метаболизма (желчные пигменты, избыток холестерола, продукты обезвреживания).
Роль печени в обмене углеводов. Основная роль печени в обмене углеводов заключается в поддержании постоянного уровня глюкозы в крови. Это осуществляется путём регуляции соотношения процессов образования и утилизации глюкозы в печени. В клетках печени содержится фермент глюкокиназа, катализирующий реакцию фосфорилирования глюкозы с образованием глюкозо-6-фосфата. Глюкозо-6-фосфат является ключевым метаболитом углеводного обмена; основные пути его превращения представлены на рисунке 1.
31.2.1. Пути утилизации глюкозы. После приёма пищи большое количество глюкозы поступает в печень по воротной вене. Эта глюкоза используется прежде всего для синтеза гликогена (схема реакций приводится на рисунке 2). Содержание гликогена в печени здоровых людей обычно составляет от 2 до 8% массы этого органа. Гликолиз и пентозофосфатный путь окисления глюкозы в печени служат в первую очередь поставщиками метаболитов-предшественников для биосинтеза аминокислот, жирных кислот, глицерола и нуклеотидов. В меньшей степени окислительные пути превращения глюкозы в печени являются источниками энергии для обеспечения биосинтетических процессов.
Рисунок 1. Главные пути превращения глюкозо-6-фосфата в печени. Цифрами обозначены: 1 - фосфорилирование глюкозы; 2 - гидролиз глюкозо-6-фосфата; 3 - синтез гликогена; 4 - мобилизация гликогена; 5 - пентозофосфатный путь; 6 - гликолиз; 7 - глюконеогенез.
Рисунок 2. Схема реакций синтеза гликогена в печени.
Рисунок 3. Схема реакций мобилизации гликогена в печени.
31.2.2. Пути образования глюкозы. В некоторых условиях (при голодании низкоуглеводной диете, длительной физической нагрузке) потребность организма в углеводах превышает то количество, которое всасывается из желудочно-кишечного тракта. В таком случае образование глюкозы осуществляется с помощью глюкозо-6-фосфатазы, катализирующей гидролиз глюкозо-6-фосфата в клетках печени. Непосредственным источником глюкозо-6-фосфата служит гликоген. Схема мобилизации гликогена представлена на рисунке 3. Мобилизация гликогена обеспечивает потребности организма человека в глюкозе на протяжении первых 12 - 24 часов голодания. В более поздние сроки основным источником глюкозы становится глюконеогенез - биосинтез из неуглеводных источников. Основными субстратами для глюконеогенеза служат лактат, глицерол и аминокислоты (за исключением лейцина). Эти соединения сначала превращаются в пируват или оксалоацетат - ключевые метаболиты глюконеогенеза. Глюконеогенез - процесс, обратный гликолизу. При этом барьеры, создаваемые необратимыми реакциями гликолиза, преодолеваются при помощи специальных ферментов, катализирующих обходные реакции (см. рисунок 4). Из других путей обмена углеводов в печени следует отметить превращение в глюкозу других пищевых моносахаридов - фруктозы и галактозы.
Рисунок 4. Гликолиз и глюконеогенез в печени. Ферменты, катализирующие необратимые реакции гликолиза: 1 - глюкокиназа; 2 - фосфофруктокиназа; 3 - пируваткиназа. Ферменты, катализирующие обходные реакции глюконеогенеза: 4 -пируваткарбоксилаза; 5 - фосфоенолпируваткарбоксикиназа; 6 -фруктозо-1, 6-дифосфатаза; 7 - глюкозо-6-фосфатаза.
Популярное:
|
Последнее изменение этой страницы: 2016-05-29; Просмотров: 1567; Нарушение авторского права страницы