
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Последовательность событий стадии элонгации
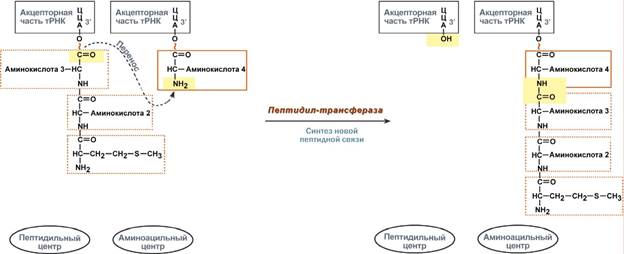
Второе повторение цикла – начинается с присоединения третьей аминоацил-тРНК к третьему кодону мРНК, аминокислота-3 становится в А-центр. Далее трансферазная реакции повторяется и образуется трипептид, занимающий А- центр, после чего он смещается в П-центр в транслоказной реакции.. В пустой А-центр входит четвертая аминоацил-тРНК и начинается третий цикл элонгации:
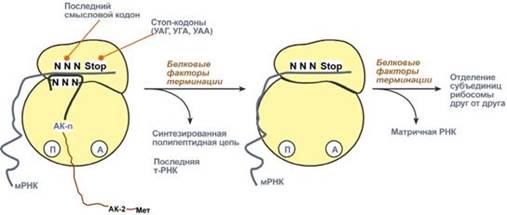
Образование пептидной связи при встраивании четвертой аминокислоты в пептид. Цикл элонгации (реакции 1, 2, 3) повторяется столько раз, сколько аминокислот необходимо включить в полипептидную цепь. Терминация Синтез белка продолжается до тех пор, пока рибосома не достигнет на мРНК особых терминирующих кодонов – стоп-кодонов УАА, УАГ, УГА. Данные триплеты не кодируют ни одной из аминокислот, их также называют нонсенс-кодоны. При вхождении этих кодонов внутрь рибосомы происходит активация белковых факторов терминации, которые последовательно катализируют: 1. Гидролитическое отщепление полипептида от конечной тРНК. 2. Отделение от П-центра последней, уже пустой, тРНК. 3. Диссоциацию рибосомы. Источником энергии для завершения трансляции является ГТФ.
Реакции стадии терминации Полирибосомы По причине того, что продолжительность жизни матричной РНК невелика, перед клеткой стоит задача использовать ее максимально эффективно, т.е. получить максимальное количество " белковых копий". Для достижения этой цели на каждой мРНК может располагаться не одна, а несколько рибосом, встающих последовательно друг за другом и синтезирующих пептидные цепи. Такие образования называются полирибосомы.
72. Посттрансляционная модификация Кратко: Посттрансляционная модификация — это ковалентная химическая модификация белка после его синтеза на рибосоме. Для многих белков посттрансляционная модификация оказывается завершающим этапом биосинтеза, который является частью процесса экспрессии генов. Полный ответ: После завершения трансляции большая часть белков подвергается дальнейшим химическим модификациям, которые называются посттрансляционными модификациями. Известно более двухсот вариантов посттрансляционных модификаций белков. Посттрансляционные модификации могут регулировать продолжительность существования белков в клетке, их ферментативную активность и взаимодействия с другими белками. В ряде случаев посттрансляционные модификации являются обязательным этапом созревания белка, в противном случае он оказывается функционально неактивным. К примеру, при созревании инсулина и некоторых других гормонов необходим ограниченный протеолиз полипептидной цепи, а при созревании белков плазматической мембраны — гликозилирование. Посттрансляционные модификации могут быть как широко распространёнными, так и редкими, вплоть до уникальных. Примером универсальной модификации служит убиквитинирование (присоединение к белку цепи из нескольких молекул короткого белка убиквитина), которое служит сигналом к расщеплению этого белка протеасомой. Другой распространённой модификацией является гликозилирование — считается, что около половины белков человека гликозилировано. К редким модификациям относят тирозинирование/детирозинирование и полиглицилирование тубулина. Один и тот же белок может подвергаться многочисленным модификациям. Так, гистоны(белки, входящие в состав хроматина у эукариот) в разных условиях могут подвергаться более чем 150 различным модификациям. Посттрансляционные модификации делят на: · модификации главной цепи; o отщепление N-концевого остатка метионина; o ограниченный протеолиз — удаление фрагмента белка, которое может происходить с концов (отщепление сигнальных последовательностей) или, в отдельных случаях, в середине молекулы (созревание инсулина); o присоединение различных химических групп к свободным амино- и карбоксильной группам (N-ацилирование, миристоилирование и др.); · модификации боковых цепей аминокислот; o присоединение или отщепление небольших химических групп (гликозилирование, фосфорилирование и др.); o присоединение липидов и углеводородов; o изменение стандартных аминокислотных остатков на нестандартные (образование цитруллина); o образование дисульфидных мостиков между остатками цистеина; · присоединение небольших белков (сумоилирование и убиквитинирование).
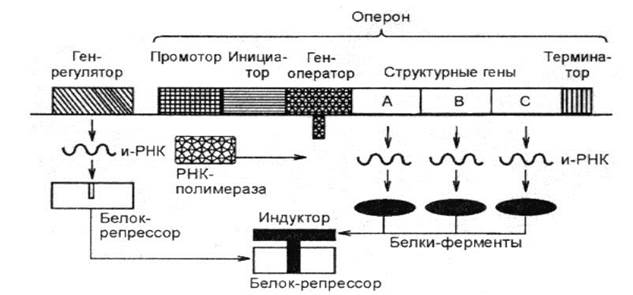
Строение оперона прокариот. Краткий обзор Основой регуляции процессов синтеза белка является оперон – комплекс генов, включающий: - ген-регулятор (Р), обеспечивающий синтез белка-репрессора (РЕП); - ген-оператор (О), управляет включением и выключением считывания информации со структурных генов, способен взаимодействовать с белком-репрессором; - ген-промотор (П) – место прикрепления РНК-полимеразы – фермента, осуществляющего процесс транскрипции; - структурные гены (СГ, содержат информацию о последовательности аминокислот в ферментах). - ген-терминатор - участок в конце оперона, сигнализирующий о прекращении транскрипции.
Механизм функционирования системы регуляции синтеза белка был открыт в 1962 году Жакобом и Моно при исследовании культивирования кишечной палочки в лактозной среде и назван lac-опероном. Упрощенно этот механизм может быть описан следующим образом. На основе информации гена-регулятора синтезируется белок-репрессор; если он активный, он связывается с геном-оператором, перекрывая путь для РНК-полимеразы – процесс трансляции и последующего синтеза белка выключается (запрещается). Если появляется индуктор (например, лактоза в lac-опероне), он присоединяется к белку-репрессору, приводя его в неактивное состояние. Оператор становится активным и включает процесс считывания информации со структурных генов – разрешает трансляцию. Происходит считывание информации с ДНК, начинается синтез необходимого белка – фермента (например, β -галактозидазы в lac-опероне).
74. Рибозимы Кратко: Рибозимы (ribozymes) [англ. ribo(se) — рибоза, от перестановки букв в англ. arabinose — арабиноза и греч. zyme — закваска, дрожжи] — природные или синтетические молекулы РНК, обладающие ферментативной активностью; в частности, рибозим катализирует расщепление и лигирование различных РНК-субстратов. Полный ответ: Рибозимы (ribozymes) [англ. ribo(se) — рибоза, от перестановки букв в англ. arabinose — арабиноза и греч. zyme — закваска, дрожжи] — природные или синтетические молекулы РНК, обладающие ферментативной активностью; в частности, рибозим катализирует расщепление и лигирование различных РНК-субстратов. Впервые ибозимы обнаружены Т. Чехом в 1980 г. при изучении аутокаталитической реакции сплайсинга предшественника рибосомной РНК уTetrahymena. За открытие рибозимов Т. Мех и С. Альтман получили Нобелевскую премию за 1989 г. Регуляция экспрессии генов с помощью антисмысловых РНК характеризуется высокой специфичностью. Это обусловлено большой точностью процесса РНК-РНК-гибридизации, основанной на комплементарном взаимодействии друг с другом протяженных последовательностей нуклеотидов. Однако сами по себе антисмысловые РНК не инактивируют необратимо мРНК-мишени, и для подавления экспрессии соответствующих генов требуются высокие (по крайней мере, эквимолярные по отношению к мРНК) внутриклеточные концентрации антисмысловых РНК. Эффективность действия антисмысловых РНК резко повысилась после того, как в их состав были введены молекулы рибозимов - коротких последовательностей РНК, обладающих эндонуклеазной активностью. Известно множество других ферментативных активностей, ассоциированных с РНК. Поэтому рибозимами в широком смысле называют молекулы РНК, обладающие любой ферментативной активностью. На модельных системах опробован РНК-овый вариант подавления ВИЧ- инфекции. Для этой цели используется необычное свойство некоторых молекул РНК - их способность разрушать другие виды РНК. Американцы Т. Чех и С. Альтман за это открытие получили в 1989 году Нобелевскую премию. Считалось, что все биохимические реакции в организме происходят благодаря высокоэффективным специфическим катализаторам, которыми служат белки - ферменты. Однако оказалось, что некоторые виды РНК, подобно белкам, обладают высокоспецифической каталитической активностью. Эти РНК назвали рибозимами. Рибозимы содержат внутри себя антисмысловые участки и участки, осуществляющие ферментативную реакцию. Т.е. они не просто присоединяются к мРНК, а еще и разрезают ее. Суть приема подавления ВИЧ-инфекции с помощью рибозимов изображена на рис. 32. Присоединяясь к комплементарной РНК-мишени, рибозим расщепляет эту РНК, результатом чего является прекращение синтеза белка, кодируемого РНК-мишенью. Если такой мишенью для рибозима будет вирусная РНК, то рибозим ее " испортит", и соответствующий вирусный белок образовываться не будет. В результате вирус прекратит свое размножение в клетке. Такой подход применим и к некоторым другим патологиям человека, например, для лечения рака.
75. РНК-интерференция Кратко: RNA interference, RNAi) — процесс подавления экспрессии гена на стадии транскрипции, трансляции, деаденилирования или деградации мРНК при помощи малых молекул РНК. Процессы РНК - интерференции обнаружены в клетках многих эукариот: у животных, растений и грибов. Полный ответ: РНК интерференция (RNA interference, RNAi): способ посттранскрипционного подавления экспрессиигенов ( сайленсинга, silencing ), при котором двухцепочечная РНК (dsRNA) индуцирует деградацию гомологичной мРНК. Двухцепочечная РНК при этом распадается на короткие двухцепочечные фрагменты (21-25 п.о.), обозначаемые как малые интерферирующие РНК (small interfering RNA, siRNA). Возможно, частично РНК интерференция может осуществляться и на транскрипционном уровне, по крайней мере, у некоторых организмов. МикроРНК (miRNA) и siRNA образуются с помощью одного и того же ферментаDICER - рибонуклеазы III типа. В отличие от miRNA, siRNA продуцируются из длинных двухцепочечных РНК, попадающих в клетку экзогенно или из dsPHK, образующихся в результате транскрипции в противоположных направлениях одного участка генома. RNAi стала важным лабораторным методом исследования функций генов в культурах клеток и в живых организмах. При этом синтетическая dsPHK, направленная против определенного транскрипта, добавляется к клеткам, чтобы вызвать RNAi На избирательное подавление синтеза вирусных белков с помощью механизма РНК-интерференции возлагаются серьезные терапевтические надежды, поскольку интерференция - существующий механизм в клетках, который мешает распространению вирусной инфекции. (Белки, участвующие в этом процессе, были названы интерферонами). В ответ на введение в клетки фрагментов нуклеиновой кислоты длиной более тридцати нуклеотидов (вирусы имеют значительно больший размер) в их цитоплазме запускается мощный интерфероновый ответ, блокирующий весь белковый синтез (т.е. происходит неспецифический защитный ответ клетки на вирусную инфекцию). И вот недавно открыт совершенно новый способ регуляции работы генов в клетках - механизм РНК-интерференции. Выяснилось, что в клетке существует специальный механизм, способный деградировать строго определенные РНК (без участия рибозимов) и таким образом полностью инактивировать её. Этот механизм деградации может быть направлен на любую конкретную РНК - клеточную, бактериальную или вирусную. Суть механизма РНК-интерференции заключается в том, что при введении в клетки короткой двунитевой РНК (днРНК) она способна вызывать специфическое разрушение той мРНК, с которой имеет гомологию. Сначала днРНК разрезается специальным ферментом на короткие фрагменты размером от 19 до 21 пар нуклеотидов. После небольших химических модификаций эти короткие днРНК образуют специфический комплекс с определенными клеточными белками. В этом комплексе днРНК расплетается и становится однонитевой. Затем короткая однонитевая РНК в силу своей комплементарности взаимодействует со строго определенной мРНК (копией гена-мишени), что является сигналом для " разрезания" последней ферментами комплекса. Образующиеся в результате этого короткие фрагменты мРНК уже не способны обеспечивать синтез полноценного белка. Таким образом, конструируя различные днРНК можно подавлять синтез строго определенных белков в клетке, не изменяя при этом структуру кодирующих их генов. Первые попытки применить РНК-интерференцию в качестве нового подхода к терапии ВИЧ-инфекции появились в 2002 г. Для того, чтобы использовать механизм РНК-интерференции, внутрь клеток нужно ввести готовые двухцепочечные молекулы РНК рис. 33. Оптимальный размер таких синтетических РНК составляет 21-28 пар нуклеотидов. Если увеличить ее длину - клетки ответят выработкой интерферона и снижением синтеза белка. Но молекулы РНК синтезировать трудно, они не очень стабильны. Поэтому на практике пользуются возможностями, предоставляемыми рекомбинантными ДНК, которые, будучи перенесенными в клетки, обеспечивают синтез таких днРНК. С целью воздействия на ВИЧ опробовано несколько вариантов РНК- интерференции. Если первоначально для подавления вируса использовали интерферирующие РНК к вирусным мРНК, то в дальнейшем пришли к выводу, что более целесообразным является направлять днРНК на клеточные мРНК, такие как, например, мРНК, кодирующие вирусный рецептор CD4 и/или корецептор CCR5. Дело в том, что вирусные гены быстро видоизменяются, соответственно изменяется нуклеотидная последовательность вирусной РНК. В результате этого конкретная искусственно синтезированная интерферирующая РНК становится мало эффективной к некоторым вариантам вируса. Клеточные же гены, обеспечивающие взаимодействие вируса с клеткой, стабильны (они изменяются чрезвычайно редко). По этой причине подавление их работы с помощью интерферирующих РНК происходит более надежно. Внимание было обращено на клеточные белки-рецепторы или корецепторы для ВИЧ. Если предотвратить синтез хотя бы одного из них, вирус не сможет проникать в клетку. На клетках, которые растут вне организма (in vitro), были использованы обе стратегии, и они дали положительный результат. Так, показано полное подавление инфицирования ВИЧ макрофагов с помощью комбинации коротких интерферирующих днРНК, направленных против клеточных и вирусных генов. Однократное применение днРНК обеспечивало долгосрочную защиту этих неделящихся клеток от вируса. Обнаружена также возможность подавления размножения ВИЧ в уже инфицированных клетках. Использование интерферирующих РНК имеет ряд преимуществ по сравнению с антисмысловыми РНК (большая эффективность, меньшая токсичность). Однако до практического применения этой новой технологии к человеку пока еще дело не дошло. Трудность заключается в том, что двунитевые РНК очень нестабильны и быстро разрушаются в организме. Кроме того, надо обеспечить присутствие этих РНК в клетках, пораженных вирусом, а не вообще в организме. На сегодняшний день еще нет надежных способов доставки генов и РНК в отдельные клетки, не отработаны до конца приемы, обеспечивающие длительное пребывание РНК в организме. Популярное:
|
Последнее изменение этой страницы: 2016-07-13; Просмотров: 3520; Нарушение авторского права страницы
 Основная часть
Основная часть