
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Физиологические типы гемоглобина.⇐ ПредыдущаяСтр 18 из 18
1. Примитивный – HbP (относятся гемоглобины, называемые Говер 1 и Говер 2) 2. Фетальный гемоглобин HbF (гемоглобин плода). 3. Гемоглобин взрослых: HbА1, HbА2, HbА3. HbР появляется на ранних стадиях развития эмбриона. Примитивные гемоглобины заменяются на HbF. На поздних стадиях развития плода появляются гемоглобины взрослых – HbА1, HbА2. В крови взрослого человека примерно 95-96% HbА1, 2-3% HbА3, 0, 1-0, 2% HbF. Гемоглобин А1 содержит по 2 Аномальные типы гемоглобина В крови человека открыто около 150 типов мутантных гемоглобинов. Аномальные гемоглобины различаются по форме, химическому составу, величине заряда. Выделены аномальные гемоглобины при помощи методов электрофореза и хроматографии. Передающиеся по наследству изменения – результат мутации единственного триплета, который приводит к замене одной аминокислоты на другую (с резко отличающимися свойствами – пример серповидноклеточная анемия – глу заменен на вал). Патология обмена гемоглобина. Болезни гемоглобинов (их около 200) называют гемоглобинозами. Гемоглобинозы делят на: 1. Гемоглобинопатии – в основе лежат наследственные изменения структуры какой-либо цепи нормального гемоглобина («молекулярные болезни»). 2. Талассемии – нарушение синтеза какой-либо цепи гемоглобина. 3. Железодефицитные анемии. Классическим примером наследственной гемоглобинпатии является серповидноклеточная анемия. Глу в 6-м положении в Талассемии – генетически обусловленной нарушение синтеза одной из нормальных цепей гемоглобина. Угнетение синтеза
Порфирии. Порфирии – группа заболеваний с наследственной предрасположенностью, возникающих в результате блокирования начальных стадий синтеза гема и сопровождающихся увеличением содержащихся порфиринов в организме.
порфобилиноген
уропорфириноген
копропорфириноген
протопорфирин IX
Гем Блок – 1 – острая перемеживающая порфирия. Накапливается Блок – 2 – приводит к накоплению всех предшествующих продуктов. Молекулярный механизм неизвестен. Болк – 3 – эритропоэтическая протопорфирия связана с отсутствием фермента феррохелатазы, каторая присоединяет к молекуле протопорфирина IX Fe2+. Распад гема За сутки в организме распадается около 9 г гемопротеидов. Период жизни эритроцитов 120 дней, разрушаются они в кровеносном русле или в селезенке. Гемоглобин связывается с гаптоглобином и в виде комплекса гаптоглобин-гемоглобин поступает в клетки ретикулоэндотелиальной системы селезенки. Комплекс гаптоглобин-гемоглобин распадается и гаптоглобин переходит в кровь, а гемоглобин окисляется в метгемоглобин (Fe3+). В РЭС селезенки гемоглобин под действием гемоксигеназы превращается в вердоглобин. Вердоглобин теряет Fe, которое связывается трансферином и доставляется кровью в костный мозг. Вердоглобин отдает белок глобин и превращается в биливердин. При восстановлении биливердина НАДФ Н2 образуется билирубин. Билирубин – плохо растворимое соединение и в крови связывается с альбумином. В виде комплекса альбумин-билирубин идет транспорт билирубина кровью в клетки печени. В печени билирубин соединяется с глюкуроновой кислотой с образованием моно (20%) и диклюкуронидов (80%), они хорошо растворимы в воде. Этот вид билирубина называется конъюгированным билирубином (связан с глюкуроновой кислотой), а также называется связанным прямым, т.к. может быть прямо обнаружен с помощью реактива Эрлиха. Билирубинглюкурониды в незначительных количествах диффундируют в кровеносный капилляр. В плазме крови присутствуют 2 формы билирубина: неконъюгированный (непрямой, свободный) и конъюгированный (прямой, связанный) – 25% от общего билирубина. Билирубинглюкурониды с желчью поступают в кишечник, где от них отщепляется глюкуроновая кислота и вновь образуется неконъюгированный билирубин. В тонком кишечнике небольшая часть билирубина может всосаться и через портальную вену вновь поступать в печень. Остальной билирубин подвергается действию кишечных бактерий и в тонком кишечнике билирубин превращается в уробилиноген. Уробилиноген всасывается в тонком кишечнике и через воротную вену поступает в печень, где уробилиноген разрушается до моно- дипирролов. Не разрушенный уробилиноген вновь с желчью поступает в кишечник и восстанавливается до стеркобилиногена (бесцветен). Стеркобилиноген окисляется до стеркобилина и выделяется с фекалиями. Небольшое количество стеркобилиногена поступает в почки, затем окисляется до стеркобилина и выделяется с мочой. В норме содержание общего билирубина в сыворотке крови составляется 8-20 мкмоль/л.
Билет 93
Синтез гема. Исходными веществами для синтеза гема служат глицин сукцинил-КоА. На первой стадии из глицина и сукцинил-КоА при участии СООН СН2-NH2 HSKoA COOH
| СН2 натсинтаза CH2 | | COSKoA C=O | CH2-NH2 Затем из 2 молекул Источником Fe2+ в синтезе гема является ферритин, который считается резервом гемопротеидом и откладывается в клетках костного мозга, печени и селезенки. Установлено, что источником всех 4 атомов азота и 8 атомов углерода тетрапиррольного кольца является глицин. Источником остальных 26 из 34 атомов углерода служит сукцинил-КоА. 4. Талассемии – нарушение синтеза какой-либо цепи гемоглобина. 5. Железодефицитные анемии. Классическим примером наследственной гемоглобинпатии является серповидноклеточная анемия. Глу в 6-м положении в Талассемии – генетически обусловленной нарушение синтеза одной из нормальных цепей гемоглобина. Угнетение синтеза
Распад гема За сутки в организме распадается около 9 г гемопротеидов. Период жизни эритроцитов 120 дней, разрушаются они в кровеносном русле или в селезенке. Гемоглобин связывается с гаптоглобином и в виде комплекса гаптоглобин-гемоглобин поступает в клетки ретикулоэндотелиальной системы селезенки. Комплекс гаптоглобин-гемоглобин распадается и гаптоглобин переходит в кровь, а гемоглобин окисляется в метгемоглобин (Fe3+). В РЭС селезенки гемоглобин под действием гемоксигеназы превращается в вердоглобин. Вердоглобин теряет Fe, которое связывается трансферином и доставляется кровью в костный мозг. Вердоглобин отдает белок глобин и превращается в биливердин. При восстановлении биливердина НАДФ Н2 образуется билирубин. Билирубин – плохо растворимое соединение и в крови связывается с альбумином. В виде комплекса альбумин-билирубин идет транспорт билирубина кровью в клетки печени. В печени билирубин соединяется с глюкуроновой кислотой с образованием моно (20%) и диклюкуронидов (80%), они хорошо растворимы в воде. Этот вид билирубина называется конъюгированным билирубином (связан с глюкуроновой кислотой), а также называется связанным прямым, т.к. может быть прямо обнаружен с помощью реактива Эрлиха. Билирубинглюкурониды в незначительных количествах диффундируют в кровеносный капилляр. В плазме крови присутствуют 2 формы билирубина: неконъюгированный (непрямой, свободный) и конъюгированный (прямой, связанный) – 25% от общего билирубина. Билирубинглюкурониды с желчью поступают в кишечник, где от них отщепляется глюкуроновая кислота и вновь образуется неконъюгированный билирубин. В тонком кишечнике небольшая часть билирубина может всосаться и через портальную вену вновь поступать в печень. Остальной билирубин подвергается действию кишечных бактерий и в тонком кишечнике билирубин превращается в уробилиноген. Уробилиноген всасывается в тонком кишечнике и через воротную вену поступает в печень, где уробилиноген разрушается до моно- дипирролов. Не разрушенный уробилиноген вновь с желчью поступает в кишечник и восстанавливается до стеркобилиногена (бесцветен). Стеркобилиноген окисляется до стеркобилина и выделяется с фекалиями. Небольшое количество стеркобилиногена поступает в почки, затем окисляется до стеркобилина и выделяется с мочой. В норме содержание общего билирубина в сыворотке крови составляется 8-20 мкмоль/л.
Билет 94 Гаптоглобин составляет 25% от всех Разновидности Нр 1-1 (азиаты) 2-1 2-2 отличаются по набору полипептидных цепей и последовательности аминокислот. Функция – гаптоглобин образует стабильный комплекс с гемоглобином, появляющимся в плазме крови в результате внутрисосудистого гемолиза эритроцитов. Гаптоглобин – гемоглобиновый комплекс поглощается клетками РЭС, где глобин и гем подвергаются распаду, а освободившееся железо используется для синтеза гема. Такой механизм предотвращает повреждение почек гемоглобином и потерю железа с мочой. Ингибитор протеаз, транспорт В12. Низкий уровень гаптоглобина приводит к гемолитической анемии. Концентрация увеличивается при воспалении, инфекции, ревматической лихорадке, метастазирующем раке. Церулоплазмин – белок голубого цвета из-за содержания меди. М.М. 150 кД, концентрация 2 – 3 г/л, синтезируется гепатоцитами. Церулоплазмин участвует в обмене меди, регулирует уровень меди в печени, участвует в обмене витамина С, адреналина, диоксифенилаланина, серотонина, мелатонина. Уменьшение содержания цероплазмина при гепатоцеребральной дистрофии (болезнь Вильсона-Коновалова) сопровождается накоплением меди в нервной ткани и печени (врожденное нарушение). Увеличение содержания церулоплазмина специфично для меланомы и шизофрении. Три белка: Трансферрин – связывает и транспортирует железо в различные ткани, регулирует концентрацию. М.М. 90 кД, концентрация – 2 – 3 г/л, синтезируется в печени и в макрофагальной системе. Имеет 2 активных центра, связывает 2 атома железа. Связывает и переносит Zn, Cu и витамин D. Трансферрин предотвращает избыточное накопление Fe3+ в тканях и потерю его с мочой. Уменьшение содержания трансферрина отмечается при гепатитах, опухолях, нефротическом синдроме. Повышение содержания связано с усиленным распадом эритроцитов.
Билет 95 АЗОТ ОСТАТОЧНЫЙ - азот небелковых соединений (мочевины, аминокислот, мочевой кислоты, креатина и креатинина, аммиака, индикана и др.), остающихся в сыворотке крови после осаждения белков. Концентрация А. о. в сыворотке крови является ценным диагностическим показателем при многих заболеваниях. А. о. определяют в надосадочной жидкости - после удаления осажденных белков сыворотки крови центрифугированием - с помощью азотометрического метода Кьельдаля в его многочисленных модификациях, колориметрических и гипобромитных методов. Метод Кьельдаля заключается в осаждении белков трихлоруксусной кислотой, минерализации надосадочной жидкости в присутствии концентрированной серной кислоты, перегонке образовавшегося аммиака и его количественном определении. Метод Кьельдаля в наст, время используется для проверки точности других методов определения азота. Для серийных определений он мало пригоден из-за своей трудоемкости.
В норме концентрация А. о. в сыворотке крови равна 20-40 мг/100 мл, или 14, 3-28, 6 ммоль/л. Увеличение содержания А. о. в крови происходит при почечной недостаточности, а также при сердечной недостаточности, злокачественных опухолях, инф. болезнях (в результате усиления распада тканевых белков и повышения содержания в крови небелковых азотсодержащих соединений). Уменьшение концентрации А. о. отмечают при беременности.
Азотемия (азот + греч. haima кровь) — избыточное содержание в крови азотсодержащих продуктов белкового обмена — см. Почечная недостаточность. В плазме присутствуют также безазотистые органические вещества: глюкоза, нейтральные жиры и липоиды (липиды).
Билет 96.
Согласно современным предтавлениям, все клеточные и внутриклеточные мембраны устроены сходным образом: основу мембраны составляет двойной молекулярный слой липидов (липидный бислой) на котором и в толще которого находятся белки ( см. рис. 1). Липидные бислои образуются амфифильными молекулами фосфолипидов и сфингомиелина в водной фазе. Амфифильными эти молекулы называют потому, что они состоят из двух частей, различных по своей растворимости в воде: полярной " головки" , обладающей высоким сродством к воде, т. е. гидрофильной, и " хвоста" , образуемого неполярными углеводородными цепями жирных кислот; эта часть молекулы обладает низким сродством к воде, т. е. гидрофобна. В состав липидов мембран входят в основном фосфолипиды, сфингомиелины и холестерин. Например, в мембранах эритроцитов человека их содержание, составляет, соответственно 36, 30 и 22 % по весу; еще 12% приходится на гликолипиды Примером амфифильной молекулы может служить молекула фосфатидилэтаноламина, структура которой показана на рис. 2. Как и другие фосфолипиды, фосфатидилэтаноламин, в химическом отношении представляет собой сложные эфиры трехатомного спирта глицерина с двумя жирными кислотами; к третьей гидроксильной группе присоединен ортофосфат, а к нему - небольшая органическая молекула, характерная для каждого вида фосфолипидов. В рассматриваемом случае это этаноламин, но могут быть также холин, инозитол, серин и некоторые другие молекулы. В состав липидного слоя мембран входят также холестерин и сфингомиелины; последние близки к фосфолипидам по химическому строению и физическим свойствами. Белки мембран принято делить на интегральные и периферические. Интегральные белки имеют обширные гидрофобные участки на поверхности и нераствориммы в воде. С липидами мембран они связаны гидрофобными взаимодействиями и частично погружены в толщу липидного бислоя, а зачастую и пронизывают бислой, оставляя на поверхности сранительно небольшие гидрофильные участки. Отделить эти белки от мембраны удается только с помощью детергентов, типа додецилсульфата или солей желчных кислот, которые разрушают липидный слой и переводят белок в растворимую форму (солюбилизируют его) образуя с ним ассоциаты. Все дальнейшие операции по очистке интегральных белков осуществляются также в присутствии детергентов. Периферические белки связаны с поверхностью липидного бислоя электростатическими силами и могут быть отмыты от мембраны солевыми растворами.
В таблице 1 перечислены функции цитоплазматических и некоторых внутриклеточных мембран. Сами по себе перегородки, разделяющие клетки на отсеки, построены из двойного слоя липидных молекул (называемого часто липидным бислоем) и практически непроницаемы для ионов и полярных молекул, растворимых в воде. Но в этот липидный бислой встроены многочисленные белковые молекулы и молекулярные комплексы, одни из которых обладают свойствами селективных (т. е. избирательных) каналов для ионов и молекул, а другие - насосов, способных активно перекачивать ионы через мембрану. Барьерные свойства мембран и работа мембранных насосов создают неравновестное распределение ионов между клеткой и внеклеточной средой, что лежит в основе процессов внутриклеточной регуляции и передачи сигналов в форме электрического импульса между клетками. Вторая функция, общая для всех мембран - это функция " монтажной платы" или матрицы, на которой располагаются в определенном порядке белки и белковые ансамбли, образующие системы переноса электронов, запасания энергии в форме АТФ, регуляции внутриклеточных процессов гормонами, поступающими извне и внутриклеточными медиаторами, узнавания других клеток и чужеродных белков, рецепции света и механических воздействий и т Гибкая и эластичная пленка, которой по существу являются все мембраны, выполняет и определенную механическую функцию, сохраняя клетку целой при умеренных механических нагрузках и нарушениях осмотического равновесия между клеткой и окружающей средой. Общие для всех мембран функции барьера для ионов и молекул и матрицы для белковых ансамблей обеспечиваются главным образом липидным бислоем, который устроен в принципе одинаково во всех мембранах. Однако набор белков индивидуален для каждого типа мембран, что позволяет мембранам участвовать в выполнении самых разных функций в различных клетках и клеточных структурах. Некоторые из этих фукнкций упомянуты в Самосборка фосфолипидных молекул в липидных везикулы в водном растворе. Каждая фосфолипидная молекула состоит из полярной группы и жирнокислотных хвостов. В водном растворе происходит самосборка мембран (справа) и замыкание мембран с образованием липидных пузырьков, называемых липосомами (слева). Резюме Биологические мембраны, наряду с цитоскелетом, формируют структуру живой клетки. Клеточная или цитоплазматическая мембрана окружает каждую клетку. Ядро окружено двумя ядерными мембранами: наружной и внутренней. Термин " мембраны" как окружающей клетку невидимой плёнки, служащей барьером между содержимым клетки и внешней средой и одновременно - полупроницаемой перегородкой, через которую могут проходить вода и некоторые растворенные в ней вещества, был впервые использован, по-видимому, ботаниками фон Молем и независимо К. фон Негели (1817-1891) в 1855 г для объясненеия явлений плазмолиза. Дальнейшее изучение осмотических явлений в растительных клетках датским ботаником Х. де Фризом (1848-1935) послужило фундаментом при создании физико-химических теорий осмотического давления и электролитической диссоциации датчанином Я. Вант-Гоффом (1852-1911) и шедским ученым С. Аррениусом (1859-1922 ). Между 1895 и 1902 годами Э. Овертон (1865-1933) измерил проницаемость клеточной мембраны для большого числа соединений и показал прямую зависимость между способностью этих соединений проникать через мембраны и их растворимостью в липидах. В 1925 году Гортер и Грендел показали, что площадь монослоя липидов, экстрагированных из мембран эритроцитов, в два раза больше суммарной площади эритроцитов. Гортер и Грендел экстрагировали липиды из гемолизированных эритроцитов ацетоном, затем выпаривали раствор на поверхности воды и измеряли площадь образовавшейся мономолекулярной пленки липидов. Вместе с тем имелись экспериментальные данные, которые свидетельствовали о том, что биологическая мембрана содержит в своем составе и белковые молекулы. Билет 97. Биохимия печени Печень самый крупный из паренхиматозных органов. Она выполняет ряд ключевых функций.
1/) 1) Принимает и распределяет вещества, поступающие в организм из пищеварительного тракта, которые приносятся с кровью по воротной вене. Эти вещества проникают в гепатоциты, подвергаются химическим превращениям и в виде промежуточных или конечных метаболитов поступают в кровь и разносятся в другие органы и ткани.
2/) 2) Служит местом образования желчи.
3/) 3) Синтезирует вещества, которые используются в других тканях.
4/) 4) Инактивирует экзогенные и эндогенные токсические вещества, а также гормоны.
Популярное:
|
Последнее изменение этой страницы: 2016-08-24; Просмотров: 1757; Нарушение авторского права страницы
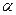 и
и 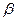 цепи. Гемоглобин А2 – по 2
цепи. Гемоглобин А2 – по 2  -цепи. Гемоглобин F – по 2
-цепи. Гемоглобин F – по 2  -цепи. Гемоглобин Говер 1 содержит 4
-цепи. Гемоглобин Говер 1 содержит 4 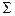 цепи, Говер 2 – 2
цепи, Говер 2 – 2  -цепями.
-цепями. Глицин Сукцинил-КоА
Глицин Сукцинил-КоА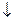


 | | ПФ |
| | ПФ | СН2 + COOH CH2 + CO2
СН2 + COOH CH2 + CO2