
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Механизм генерации потенциала действия. Распространение потенциала действия по миелиновым и безмиелиновым нервным волокнам.
Все клетки возбудимых тканей при действии различных раздражителей достаточной силы способны переходить в состояние возбуждения. Обязательным признаком возбуждения является изменение электрического состояния клеточной мембраны. Общее изменение разности потенциалов между клеткой и средой, происходящее при пороговом и сверхпороговом возбуждении клеток, называется потенциалом действия. Потенциалы действия обеспечивают проведение возбуждения по нервным волокнам и инициируют процессы сокращения мышечных и секреции железистых клеток. На основе обобщения большого экспериментального материала было установлено, что потенциалы действия возникают в результате избыточного по сравнению с покоем диффузии ионов натрия из окружающей жидкости внутрь клетки. Формирование потенциала действия обусловлено двумя ионными потоками через мембрану: поток ионов натрия внутрь клетки приводит к перезарядке мембраны, а противоположно направленный поток ионов калия обусловливает восстановление исходного потенциала покоя. Потоки приблизительно равны по величине, но сдвинуты во времени. Благодаря этому сдвигу во времени и возможно появление потенциала действия. Потенциал действия, возникнув в одном участке нервной клетки, быстро распространяется по всей ее поверхности. Распространение потенциала действия обусловлено возникновением так называемых локальных токов, циркулирующих между возбужденным и невозбужденным участками клетки. В состоянии покоя внешняя поверхность клеточной мембраны имеет положительный потенциал, а внутренняя - отрицательный. В момент возбуждения полярность мембраны изменяется на обратную: ее внешняя поверхность заряжена отрицательно по отношению к внутренней (рис.2).
Рис.2. Мембранные потенциалы.
Рис..2. Мембранные потенциалы. Рис.2. Мембранные потенциалы.
В результате этого между возбужденным и невозбужденным участками мембраны имеется разность потенциалов. Наличие разности потенциалов приводит к появлению между этими участками электрических токов, называемых локальными токами или токами действия. На поверхности клетки локальный ток течет от невозбужденного участка к возбужденному; внутри клетки он течет в обратном направлении. Локальный ток, как и любой электрический ток, оказывает раздражающее действие на соседние невозбужденные участки и вызывает увеличение проницаемости их мембран. Это приводит к снижению в них потенциала покоя. Когда деполяризация достигает критического значения, в этих участках возникают потенциалы действия, а в том участке, который ранее был возбужденным, в это время уже происходят восстановительные процессы реполяризации. Вновь возбужденный участок, в свою очередь, становится электроотрицательным и возникающий локальный ток раздражает следующий за ним участок. Этот процесс многократно повторяется и обусловливает распространение импульсов возбуждения по всей длине клетки в обоих направлениях. В нервной системе прохождение импульсов только в определенном направлении обусловлено наличием синапсов, обладающих односторонней проводимостью. Под влиянием локальных токов волна возбуждения распространяется вдоль волокна без затухания (бездекрементное проведение). Это обусловлено тем, что локальные токи только деполяризуют мембрану до критического уровня, а потенциалы действия в каждом участке мембраны поддерживаются независимыми ионными потоками, перпендикулярными к направлению распространения возбуждения. Скорость уменьшения мембранного потенциала до критического уровня зависит от разности потенциалов между возбужденным и невозбужденным участками и от кабельных свойств волокна: электрической емкости и сопротивления мембраны, сопротивлений аксоплазмы и окружающей среды. Кабельные свойства волокна обеспечивают деполяризацию мембраны до критического уровня, а последующая диффузия натрия в клетку усиливает деполяризацию и обеспечивает незатухающее проведение импульса. Количественно процесс может быть представлен следующим образом. Скорость распространения потенциала действия определяется в основном тем временем, которое необходимо для критической деполяризации мембраны. Обозначим деполяризацию до критического уровня " Dj ". Тогда " Dj ", емкость мембраны " с" и заряд " Dq", необходимый для деполяризации мембраны на " Dj " будут:
Заряд мембраны и ее потенциал изменяется вследствие протекания локального тока " I". Тогда время " t", в течение которого заряд мембраны изменяется на Dq, а потенциал на " Dj " будет равно:
Из уравнений (4) и (5) получим:
Величина локального тока " I" по закону Ома определяется разностью потенциалов " U" между возбужденным и невозбужденным участками мембраны и сопротивлением " R" всех участков, по которым протекает локальный ток: В основном оно определяется сопротивлением мембраны. Подставляя значение " I " в уравнение (6), получим:
Учитывая, что " Dj " и " U" величины постоянные, можно написать:
где k - коэффициент пропорциональности. Уравнение (7) показывает, что время деполяризации мембраны и, следовательно, время проведения возбуждения пропорциональны произведению " RC", называемому постоянной времени мембраны. Тогда скорость проведения возбуждения будет обратно пропорциональна постоянной времени мембраны. Скорость проведения импульса возрастает с увеличением диаметра волокна. Это объясняется тем, что с увеличением диаметра уменьшается сопротивление, приходящееся на единицу длины волокна. В нервных волокнах характер распространения возбуждения зависит от наличия или отсутствия в них миелиновых оболочек. В безмякотных волокнах возбуждение распространяется непрерывно вдоль всей мембраны. Все участки мембраны при этом в свое время становятся возбужденными. В мякотных нервных волокнах возбуждение распространяется несколько по иному. Мякотные нервные волокна имеют толстые миелиновые оболочки, которые через 1 - 3 мм прерывается с образованием так называемых перехватов Ранвье.
Рис.3. Нервные волокна с толстыми миелиновыми оболочками.
В электрическом отношении миелин является изолятором; его удельное сопротивление в 10 млн. раз превышает удельное сопротивление раствора Рингера. В результате этого локальные токи через миелиновые оболочки протекать не могут; они циркулируют между перехватами Ранвье. При возбуждении одного перехвата Ранвье между ним и следующим перехватом возникают локальные токи и импульс как бы перескакивает на второй перехват, со второго - на третий и т.д. Такой способ проведения нервного импульса называется сальтаторным. При блокировании одного перехвата Ранвье каким-либо анестезирующим веществом, например кокаином, импульс сразу передается на третий перехват. При блокировании сразу двух перехватов импульс дальше распространяться не может: сопротивление между первым и четвертым перехватами велико, и локальный ток между ними не достигает порогового значения. В результате сальтаторного способа передачи скорость распространения первого импульса в мякотных волокнах примерно в 10 раз выше, чем в безмякотных, при одинаковом диаметре волокон. Помимо этого, сальтаторный способ проведения возбуждения является более экономичным, поскольку в этом случае ионные потоки проходят не через всю поверхность клетки, как при непрерывном распространении, а только через поверхность в области перехватов Ранвье.
1.3. Основные характеристики электрического поля. Электрический диполь. Поле диполя. Диполь в электрическом поле.
Электрическим диполем (диполем) называют систему, состоящую из двух равных, но противоположных по знаку точечных электрических зарядов, расположенных на некотором расстоянии друг от друга (плечо диполя). Основной характеристикой диполя (рис.4) является его электрический, или дипольный момент – вектор, направленный от отрицательного заряда к положительному и равный произведению заряда на плечо диполя:
Рис.4. Электрический диполь.
Единицей электрического момента диполя является кулон-метр. Поместим диполь в однородное электрическое поле напряжённостью
M=qEl sina=pEsina, (9)
Или в векторной форме
Рис.5. Диполь в однородном электрическом поле.
Таким образом, на диполь в однородном электрическом поле действует вращающий момент, зависящий от электрического момента, ориентации диполя в поле и напряжённости поля. Рассмотрим теперь диполь в неоднородном электрическом поле. Простоты ради предположим, что диполь расположен вдоль силовой линии. На него действуют силы
где
F= F_ - F+ = qE_ - qE+ = q (E_ - E+). (11)
Рис.6. Силы, действующие на диполь. Введём отношение (E_ - E+)/l, характеризующее среднее изменение напряжённости, приходящееся на единицу длины диполя. Так как обычно плечо невелико, то приближенно можно считать
(Е_ - Е+)/l = dE/dx, (12) где dE/dx – производная от напряженности электрического поля по направлению оси ОХ, являющаяся мерой неоднородности электрического поля вдоль соответствующего направления. Из (12) следует
Е_ - E+ = l dE/dx, Тогда формулу (5) можно представить в виде
F = ql dE/dx = p dE/dx. Итак, на диполь действует сила, зависящая от его электрического момента и степени неоднородности поля dE/dx. Если диполь ориентирован в неоднородном электрическом поле не вдоль силовой линии, то на него дополнительно действует еще и вращающий момент. Так что свободный диполь практически всегда будет втягиваться в область больших значений напряженности поля. Популярное:
|
Последнее изменение этой страницы: 2016-08-31; Просмотров: 746; Нарушение авторского права страницы

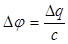 (4)
(4) (5)
(5) (6)
(6)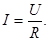 Сопротивление " R" суммируется из сопротивлений всех последовательных участков, по которым протекает локальный ток: мембраны, цитоплазмы, окружающей клетку жидкости.
Сопротивление " R" суммируется из сопротивлений всех последовательных участков, по которым протекает локальный ток: мембраны, цитоплазмы, окружающей клетку жидкости.
 (7)
(7)
 (8)
(8)
 (рис.5). На каждый из зарядов диполя действуют силы
(рис.5). На каждый из зарядов диполя действуют силы  =q
=q  = -q
= -q 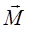 =
=  (10)
(10)
 +=
+=  + и
+ и 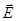 + и
+ и 