|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Прокариотная клетка. Особенности строенияСтр 1 из 5Следующая ⇒
Прокариотная клетка. Особенности строения Строение прокариотной клетки Деления клеточных организмов на высшем уровне предусматривают выделение всех прокариот в отдельное царство. В 70-х гг. обнаружены микроорганизмы, структурно относящиеся к прокариотному типу, но значительно отличающиеся химическим строением важных клеточных макромолекул и способностью осуществлять уникальные биохимические процессы. Эти необычные прокариотные организмы были названы архебактериями. Типичные прокариоты, или бактерии, получили соответственно название эубактерий (истинных бактерий). Число известных архебактерий по сравнению с эубактериями чрезвычайно мало. Материал настоящей работы посвящен общей характеристике прокариотных организмов (в основном эубактерий), отличающихся морфологическим и особенно физиологическим разнообразием. В основе морфологического разнообразия лежат различия в размерах и форме отдельных клеток, способах их деления, природе и наборе цитоплазматических включений, строении клеточной стенки и структур, локализованных снаружи от нее, наличии и типе дифференцированных форм, образующихся в процессе жизненного цикла. Форма прокариот
Рис. 1 Разнообразие форм прокариот 1 — кокк; 2 — диплококк; 3 — сарцина; 4 — стрептококк; 5 — колония сферической формы: 6 — палочковидные бактерии (одиночная клетка и цепочка клеток); 7 — спириллы; 8 — вибрион; 9 — бактерии, имеющие форму замкнутого или незамкнутого кольца; 10 — бактерии, образующие выросты (простеки); 11 — бактерия червеобразной формы; 12 — бактериальная клетка в форме шестиугольной звезды; 13 — представитель актиномицетов; 14 — плодовое тело миксобактерии; 15 — нитчатая бактерия рода Caryophanon с латерально расположенными жгутиками: 16 — нитчатая цианобактерия. образующая споры (акинеты) и гетероцисты; 8, 15, 17, 18 — бактерии с разными типами жгутикования; 19 — бактерии, образующая капсулу; 20 — нитчатые бактерии группы Sphaeroillus, заключенные в чехол, инкрустированный гидратом окиси железа; 21 — бактерия, образующая шипы; 22 — Galionella. До недавнего времени большинство исследователей традиционно считали, что клетки прокариот достаточно однообразны и в подавляющем большинстве имеют форму сферы, цилиндра или спирали. Они бывают одиночными, в иных случаях образуют нити или колонии. Прокариоты сферической формы, называемые кокками, могут после деления не расходиться. Если деление происходит в одной плоскости, образуются пары клеток (диплококки) или цепочки (стрептококки). В том случае, когда деление происходит относительно равномерно в трех взаимно перпендикулярных направлениях, и клетки после деления остаются соединенными друг с другом, возникают пакеты правильной формы (сарцины) или колонии сферической формы. Если же деление происходит в нескольких плоскостях неравномерно, образуются клеточные скопления неправильной формы (рис. 1; 1 – 5). Прокариоты, имеющие форму цилиндра (палочковидные), сильно различаются по величине отношения длины клетки к ее поперечнику. Прокариоты спиралевидной формы характеризуются разным числом витков: у спирилл — от одного до нескольких витков, вибрионы выглядят наподобие изогнутых палочек, так что их можно рассматривать как неполный виток спирали (рис. 1; 6 – 8). За последнее время среди прокариот обнаружены организмы, отличающиеся от описанных выше основных форм. Некоторые бактерии имеют вид кольца, замкнутого или разомкнутого в зависимости от стадии роста (рис. 1; 9). У прокариот, в основном размножающихся почкованием, описано образование клеточных выростов (простек), число которых может колебаться от 1 до 8 и более (рис. 1; 10). Из природных субстратов выделены бактерии червеобразной формы и напоминающие шестиугольную звезду (рис. 1; 11, 12). Для некоторых видов характерно слабое или довольно хорошо выраженное ветвление (рис. 1; 13). Описаны прокариоты, обладающие морфологической изменчивостью, в зависимости от условий имеющие вид палочек, кокков или обнаруживающие слабое ветвление. Форма многоклеточных прокариот также разнообразна: это скопления различной конфигурации, чаще — нити (рис. 1; 14 – 16). Своеобразие бактериальным клеткам придают жгутики, имеющие различное расположение на клеточной поверхности (рис. 1; 8, 15, 17, 18), а также выделения внеклеточных веществ разной химической природы (рис. 1; 19 – 22). Клеточная стенка Клеточная стенка — важный и обязательный структурный элемент подавляющего большинства прокариотных клеток, располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с окружающей средой (у клеток, не содержащих этих слоев клеточной оболочки). На долю клеточной стенки приходится от 5 до 50 % сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. Концентрация солей в клетке, как правило, намного выше, чем в окружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка чисто механически защищает клетку от проникновения в нее избытка воды. По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты, относящиеся к эубактериям, делятся на две большие группы. Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна. У так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются. У некоторых эубактерий положительная реакция при окрашивании описанным выше способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки.
Рис. 3 Клеточная стенка грамположительных (А) и грамотрицательных (Б) эубактерий 1 — цитоплазматическая мембрана; 2 — пептидогликан; 3 — периплазматическое пространство; 4 — наружная мембрана: 5 — цитоплазма, в центре которой расположена ДНК. Клеточные стенки грамположительных и грамотрицательных эубактерий резко различаются как по химическому составу, так и по ультраструктуре (рис. 3). В состав клеточной стенки эубактерий входят семь различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке. У грамположительных 1 эубактерий он составляет основную массу вещества клеточной стенки (от 40 до 90 %), у грамотрицательных — содержание пептидогликана значительно меньше (1 – 10 %). Клеточная стенка цианобактерий, сходная с таковой грамотрицательных эубактерий, содержит от 20 до 50 % этого гетерополимера. Этот способ был впервые предложен в 1884 г. датским ученым X. Грамом (Ch. Gram), занимавшимся окрашиванием тканей. Позднее он был использован для бактерий. Под электронным микроскопом клеточная стенка грамположительных эубактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм. У грамотрицательных эубактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2 – 3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8 – 10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных эубактерий получил название наружной мембраны.
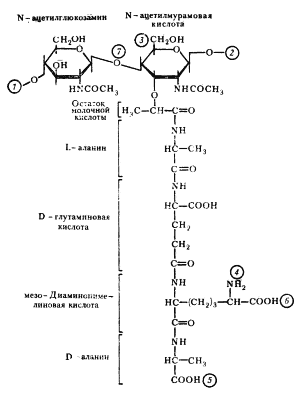
Рис. 4 Структура повторяющейся единицы пептидогликана клеточной стенки эубактерий 1, 2 — места полимеризации гликанового остова молекулы; 3 — место присоединения с помощью фосфодиэфирной связи молекулы тейхоевой кислоты в клеточной стенке грамположительных эубактерий; 4, 5 — места, по которым происходит связывание между гликановыми цепями с помощью пептидных связей; 6 — место ковалентного связывания (пептидная связь) с липопротеином наружной мембраны у грамотрицательных эубактерий; 7 — место действия лизоцима.
Клеточная стенка грамположительных эубактерий плотно прилегает к ЦПМ, в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электронно-прозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической: и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп эубактерий, характерно только для грамотрицательных форм. Клеточная стенка грамположительных эубактерий. Основную массу клеточной стенки грамположительных эубактерий составляет специфический гетерополимер — пептидогликан. Полисахаридный остов молекулы построен из чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных между собой посредством b-1, 4-гликозидных связей (рис. 4). К N-ацетилмурамовой кислоте присоединен короткий пептидный хвост, состоящий из небольшого числа (обычно 4 – 5) аминокислот. У грамположительных эубактерий обнаружено более 100 различных химических типов пептидогликана. Большинство различий относится к пептидной части его молекулы. Две особенности пептидного хвоста заслуживают внимания: наличие аминокислот в D-форме (неприродная конфигурация) и высокое содержание аминокислот с двумя аминогруппами. Это имеет принципиальное значение для пространственной организации пептидогликана. Обе аминогруппы этих аминокислот могут участвовать в образовании пептидных связей, причем вторые аминогруппы — в формировании дополнительных пептидных связей между гетерополимерными цепочками.
Рис. 5 Пептидные мостики между гетерополимерными цепочками Г — N-ацетилглюкозамин; М — N-ацетилмурамовая кислота; ала — аланин; глу — глутаминовая кислота; лиз — лизин; ДАП — диаминопимелиновая кислота; гли — глицин; стрелками обозначено место действия пенициллина. В большинстве случаев в образовании пептидной связи участвует карбоксильная группа D-аланина одного тетрапептида и свободная аминогруппа диаминокислоты другого (рис. 5, А). Иногда связь между тетрапептидами разных гликановых цепей осуществляется с помощью других аминокислот (рис. 5, Б). Нетрудно себе представить, что этим способом можно " сшить" между собой множество гетерополимерных цепей. Частота " сшивок" различна, поскольку не все пептидные хвосты участвуют в формировании межцепочечных связей. Некоторые образуют ковалентные связи с другими химическими молекулами, входящими в состав клеточной стенки, и, наконец, часть тетрапептидных хвостов находится в свободном состоянии. Пептидогликан, окружающий протопласт грамположительных эубактерий, — это по существу одна гигантская молекула, " сшитая" с помощью гликозидных и пептидных связей. Именно последние обеспечивают ей трехмерную пространственную организацию. Кроме пептидогликана в состав клеточных стенок грамположительных эубактерий входит другой уникальный класс химических соединений — тейхоевые кислоты, представляющие собой полимеры, построенные на основе рибита (пятиатомного спирта) или глицерина (трехатомного спирта), остатки которых соединены между собой фосфодиэфирными связями (рис. 6). Некоторые свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками D-аланина, глюкозы, N-ацетилглюкозамина и некоторых других сахаров. Тейхоевые кислоты ковалентно могут соединяться с N-ацетилмурамовой кислотой (см. рис. 4). Поскольку это длинные линейные молекулы, они могут пронизывать весь пептидогликановый слой, достигая внешней поверхности клеточной стенки. В этом случае, вероятно, они являются основными антигенами грамположительных эубактерий. Остающиеся свободные гидроксилы фосфорной кислоты придают тейхоевой кислоте свойства полианиона. Как полианионы тейхоевые кислоты определяют поверхностный заряд клетки. Сахарные компоненты тейхоевых кислот входят в состав рецепторов для некоторых бактериофагов и определяют возможность адсорбции фага на клеточной поверхности.
Рис. 6 Структурная формула глицеринтейхоевой кислоты Содержит чередующиеся остатки D-аланина и N-ацетилглюкозамина (по Rose, 1971). В составе клеточной стенки грамположительных эубактерий в небольших количествах также найдены полисахариды, белки и липиды. Для полисахаридов и липидов показана возможность ковалентного связывания с макромолекулами клеточной стенки, в отличие от белков, которые (у тех видов, где имеются) формируют на ее внешней поверхности отдельный слой. Таким образом, основными компонентами клеточной стенки грамположительных эубактерий являются три типа макромолекул: пептидогликаны, тейхоевые кислоты и полисахариды, которые с помощью ковалентных связей образуют сложную структуру с весьма упорядоченной пространственной организацией. Клеточная стенка бацилл, например Bacillus subtilis, приблизительно соответствует толщине 40 молекул пептидогликана. В целом клеточную стенку грамположительных эубактерий можно представить в виде губчатой структуры с порами диаметром примерно 1 – 6 нм. Возможность прохождения молекул через такую клеточную стенку определяется ее зарядом и размером пор. Клеточная стенка грамотрицательных эубактерий. У грамотрицательных эубактерий строение клеточной стенки намного сложнее, чем у грамположительных (см. рис. 3). В ее состав входит гораздо большее число макромолекул разного химического типа (см. табл. 1). Таблица 1/3 Химический состав клеточных стенок грамположительных и грамотрицательных эубактерий (по Rose, 1971; Freer, Salton, 1971)
бозначения: (–) — отсутствуют, (+) — присутствуют, (±) — присутствуют не у всех видов Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов грамотрицательных эубактерий содержание этого гетерополимера колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями (рис. 7).
Рис. 7 Однослойная структура пептидогликана Линиями обозначены гетерополимерные цепочки, образованные чередующимися остатками N-ацетилглюкозамина (Г) и N-ацетилмурамовой кислоты (М), соединенными между собой b-1, 4-гликозидными связями. Кружочками обозначены аминокислоты пептидного хвоста. Химическая структура пептидогликана грамотрицательных эубактерий в основном сходна со структурой типичного пептидогликана грамположительных эубактерий (см. рис. 4; 5, А). Снаружи от пептидогликана располагается дополнительный слой клеточной стенки — наружная мембрана. Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида (рис. 8, А). Специфическим компонентом наружной мембраны является липополисахарид сложного молекулярного строения, занимающий около 30 – 40 % ее поверхности и локализованный во внешнем слое (рис. 8, 5).
Рис. 8 А. Клеточная стенка грамотрицательных эубактерий: 1 — цитоплазматическая мембрана; 2 — пептидогликановый слой; 3 — периплазматическое пространство; 4 — молекулы белков (заштрихована гидрофобная часть); 5 — фосфолипид; 6 — липополисахарид. Б. Строение молекулы липополисахарида: 1 — липид А; 2 — внутреннее полисахаридное ядро; 3 — наружное полисахаридное ядро; 4 — О-антиген. Белки наружной мембраны можно разделить на основные и минорные. Основные белки представлены небольшим числом различных видов, но составляют почти 80 % всех белков наружной мембраны. Одна из функций этих белков — формирование в мембране гидрофильных пор диаметром примерно 1 нм, через которые осуществляется неспецифическая диффузия молекул с массой до 600 – 900 Да7 (7Да — дальтон, или единица атомной массы, равен 1, 66033 x 10-27 кг). Это означает, что через такие поры могут проходить сахара, аминокислоты, небольшие олигосахариды и пептиды. Белки, пронизывающие наружную мембрану насквозь и образующие гидрофильные поры, называют поринами. Минорные белки наружной мембраны представлены гораздо большим числом видов. Их основная функция — транспортная и рецепторная. Примером минорных белков могут служить белки, ответственные за специфический транспорт в клетку железосодержащих соединений. Помимо слоев клеточной стенки, типичных для большинства грамотрицательных эубактерий, у некоторых представителей этой группы обнаружены дополнительные слои разной электронной плотности, располагающиеся с внешней стороны от наружной клеточной мембраны. Однако до настоящего времени не ясно, относятся ли они к клеточной стенке, являясь результатом ее последующего усложнения, или же представляют собой структурные элементы многослойного чехла. Ворсинки К поверхностным структурам бактериальной клетки относятся также ворсинки (фимбрии, пили) (см. рис. 2; 6). Их насчитывается от нескольких единиц до нескольких тысяч на клетку. Эти структуры не имеют отношения к движению бактерий и обнаружены у подвижных и неподвижных форм. Ворсинки построены из одного вида белка — пилина — и представляют собой прямые белковые цилиндры, отходящие от поверхности клетки. Они, как правило, тоньше жгутиков (диаметр — 5 – 10 нм, длина 0, 2 – 2, 0 мкм), расположены перитрихиально или полярно. Больше всего сведений имеется о ворсинках Е. coli. У этой бактерии описаны ворсинки общего типа и половые. Ворсинки общего типа придают бактериям свойство гидрофобности, обеспечивают их прикрепление к клеткам растений, грибов и неорганическим частицам, принимают участие в транспорте метаболитов. Через ворсинки в клетку могут проникать вирусы. Наиболее хорошо изучены половые ворсинки, или F-пили, принимающие участие в половом процессе бактерий. F-пили необходимы клетке-донору для обеспечения контакта между ней и реципиентом и в качестве конъюгационного тоннеля, по которому происходит передача ДНК. Ворсинки нельзя считать обязательной клеточной структурой, так как и без них бактерии хорошо растут и размножаются. Мембраны Содержимое клетки отделяется от клеточной стенки цитоплазматической мембраной (ЦПМ) — обязательным структурным элементом любой клетки, нарушение целостности которого приводит к потере клеткой жизнеспособности. На долю ЦПМ приходится 8 – 15 % сухого вещества клеток. У большинства прокариотных клеток ЦПМ — единственная мембрана. В клетках фототрофных и ряда хемотрофных прокариот содержатся также мембранные структуры, располагающиеся в цитоплазме и получившие название внутрицитоплазматических мембран. Их происхождение и функции будут рассмотрены ниже. Химический состав мембран ЦПМ — белково-липидный комплекс, в котором белки составляют 50 – 75 %, липиды — от 15 до 45 %. Кроме того, в составе мембран обнаружено небольшое количество углеводов. Как правило, липиды и белки составляют 95 % и больше вещества мембран. Главным липидным компонентом бактериальных мембран являются фосфолипиды — производные 3-фосфоглицерина. Хотя у прокариот найдено множество различных фосфолипидов, набор их в значительной степени родо- и даже видоспецифичен. Широко представлены в бактериальных мембранах различные гликолипиды. Стерины отсутствуют у подавляющего большинства прокариот, за исключением представителей группы микоплазм и некоторых бактерий. Так, в ЦПМ Acholeplasma содержится 10 – 30 % холестерина, поглощаемого из внешней среды, от общего содержания мембранных липидов. Из других групп липидов в мембранах прокариот обнаружены каротиноиды, хиноны, углеводороды.
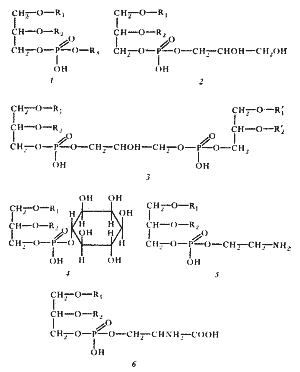
Рис. 12 Структура основных фосфолипидов мембран эубактерий. R1 и R2 — остатки длинноцепочечных жирных кислот, образующих гидрофобный " хвост" молекулы; R3 может быть остатком глицерина, его производных, этаноламина, инозита и других соединений. Эта часть составляет гидрофильную " голову" молекулы. Простейшим фосфолипидом является фосфатидная кислота, не имеющая R3-остатка, связанного с фосфорной кислотой сложноэфирной связью. 1 — общая структура фосфолипида; 2 — фосфатидилглицерин; 3 — дифосфатидилглицерин (кардиолипин); 4 — фосфатидилинозит; 5 — фосфатидилэтаноламин; 6 — фосфатидилсерин. Все липиды эубактерий — производные глицерина — содержат один или несколько остатков жирных кислот, состав которых весьма своеобразен (рис. 12). В основном это насыщенные или мононенасыщенные жирные кислоты с 16 – 18 углеродными атомами. Полиненасыщенные жирные кислоты у эубактерий отсутствуют. Исключение составляют цианобактерии, у разных видов которых найдены полиненасыщенные жирные кислоты типа C 16: 2, C 18: 2, C 18: 3, C 15: 4. Помимо обычных жирных кислот, т. е. обнаруживаемых и в клетках эукариот, в составе мембранных липидов эубактерий находят и кислоты, не встречающиеся, как правило, в мембранах эукариот. Это циклопропановые жирные кислоты, содержащие одно или больше трехчленных колец, присоединенных вдоль углеводородной цепи. Другие, редко встречающиеся и обнаруженные практически только у эубактерий кислоты — это разветвленные жирные кислоты с 15 – 17 углеродными атомами. Набор жирных кислот в мембранных липидах также чрезвычайно видоспецифичен. У некоторых грамположительных эубактерий C 15-жирная кислота с разветвленной цепью может составлять до 90 % всех жирных кислот липидов. Главная функция липидов — поддержание механической стабильности мембраны и придание ей гидрофобных свойств. Особый состав липидов обнаружен в мембранах архебактерий. У них не найдены типичные для эубактерий эфиры глицерина и жирных кислот, но присутствуют эфиры глицерина и высокомолекулярных С 20-, С 40-спиртов, а также нейтральные изопреноидные С 20 – С 30-углеводороды. На долю белков приходится больше половины сухой массы мембран. К мембранам с наиболее высоким содержанием белка относятся бактериальные ЦПМ. При изучении их белкового состава не было обнаружено какого-либо универсального структурного белка. ЦПМ Escherichia coli содержит 27 основных и множество минорных белков, но ни один из основных белков не присутствует в преобладающих количествах. Поскольку ЦПМ прокариот многофункциональна и участвует в осуществлении разнообразных ферментативных процессов, был сделан вывод, что мембранные белки — это, как правило, ферменты. По аминокислотному составу мембранные белки не отличаются от других клеточных белков, за исключением того, что в них содержится мало (иногда следы) цистеина. В некоторых бактериальных мембранах в значительных количествах обнаружены углеводы. По-видимому, они содержатся не в свободном состоянии, а входят в состав гликолипидов и гликопротеинов. Структура мембран Мембранные липиды всех эубактерий и части архебактерий образуют бислои, в которых гидрофильные " головы" молекул обращены наружу, а гидрофобные " хвосты" погружены в толщу мембраны (рис. 13). Углеводородные цепи, прилегающие к гидрофильным " головам", довольно жестко фиксированы, а более удаленные части " хвостов" обладают достаточной гибкостью. У некоторых архебактерий (ряд метаногенов, термоацидофилы) мембранные липиды, в состав которых входит C 40-спирт, формируют монослойную мембрану, по толщине равную бислойной. Монослойные липидные мембраны обладают большей жесткостью сравнительно с бислойной. При " биологических" температурах мембранные липиды находятся в жидкостно-кристаллическом состоянии, характеризующемся частичной упорядоченностью структуры. При понижении температуры они переходят в квазикристаллическое состояние. Чем более не насыщены и разветвлены остатки жирных кислот или чем большее число циклических группировок они содержат, тем ниже температура перехода из жидкостно-кристаллического состояния в квазикристаллическое. " Жидкая" структура мембран обеспечивает определенную свободу молекул белков, что является необходимым для осуществления процессов транспорта электронов и веществ через мембрану. Это же свойство обусловливает высокую эластичность мембран: они легко сливаются друг с другом, растягиваются и сжимаются.
Рис. 13 Модель строения элементарной биологической мембраны 1 — молекулы липидов: а — гидрофильная " голова"; б — гидрофобный " хвост"; 2 — молекулы белков: в — интегральная; г — периферическая; д — поверхностная. В отличие от липидов у мембранных белков нет единого способа структурной организации. 30 – 50 % белка имеет конфигурацию a-спирали, остальная часть находится преимущественно в виде беспорядочного клубка. Вероятно, часть белков лишена ферментативной активности и участвует только в поддержании мембранной структуры. В то же время доказано, что для осуществления белками некоторых функций необходима их строго упорядоченная взаимная организация в мембране. В зависимости от расположения в мембране и характера связи с липидным слоем мембранные белки условно можно разделить на три группы: интегральные, периферические и поверхностные (см. рис. 13). Интегральные белки полностью погружены в мембрану, а иногда пронизывают ее насквозь. Связь интегральных белков с мембранными липидами очень прочна и определяется главным образом гидрофобными взаимодействиями. Периферические белки частично погружены в гидрофобную область, а поверхностные находятся вне ее. В первом случае связь с липидами в основном, а во втором — исключительно определяется электростатическими взаимодействиями. Помимо этого некоторые белки и липиды в мембране могут быть связаны ковалентно. Предложено несколько моделей строения мембраны. Наибольшее признание получила модель, учитывающая большинство данных, известных о мембранах, согласно которой в липидную основу включены асимметрично расположенные белковые молекулы (см. рис. 13). Некоторые из них образуют скопления на поверхностях липидного би- или монослоя, другие частично или полностью погружены в него, третьи пронизывают его насквозь. В модели подчеркнута асимметрия строения мембраны, основанная на различиях в химическом строении и расположении молекул белка. Функции ЦПМ прокариот ЦПМ прокариот выполняет разнообразные функции, в основном обеспечиваемые локализованными в ней соответствующими ферментными белками. Первоначально была постулирована барьерная функция клеточной мембраны, получившая позднее экспериментальное подтверждение. С помощью специальных переносчиков, называемых транслоказами, через мембрану осуществляется избирательный перенос различных органических и неорганических молекул и ионов. В ней локализованы ферменты, катализирующие конечные этапы синтеза мембранных липидов, компонентов клеточной стенки и некоторых других веществ. Общепризнана роль ЦПМ прокариот в превращениях клеточной энергии. У бактерий, источником энергии для которых служат процессы дыхания или фотосинтеза, в ЦПМ определенным образом расположены переносчики цепи электронного транспорта, функционирование которых приводит к генерированию электрохимической энергии (DmH+), используемой затем в клетке по разным каналам, в том числе и для образования химической энергии (АТФ). ЦПМ является одним из компонентов аппарата генерирования DmH+. В мембране расположены также ферментные комплексы, обеспечивающие превращения: DmH+ ® АТФ. ЦПМ принимает участие в репликации и последующем разделении хромосомы прокариотной клетки. В последнее время выявляется еще одна функциональная грань клеточных мембран — их интегрирующая роль в организме, вполне сочетающаяся с давно установленной разъединяющей (барьерной) функцией. Клетка — единое целое. В обеспечении этого принципа клеточной организации важная роль принадлежит мембранам. Показан перенос электрохимической энергии и электронов вдоль мембран. Последние рассматриваются так же как возможные пути транспорта жирорастворимых субстратов и молекулярного кислорода. ЦПМ является основным барьером, обеспечивающим избирательное поступление в клетку и выход из нее разнообразных веществ и ионов. Осуществляется это с использованием разных механизмов мембранного транспорта. Выделяют 4 типа транспортных систем, с участием которых происходит проникновение молекул в бактериальную клетку: пассивную диффузию, облегченную диффузию, активный транспорт и перенос химически модифицированных молекул. У грамположительных форм ЦПМ является и единственным барьером такого рода, у грамотрицательных эубактерий функции дополнительного барьера (молекулярного " сита" ) выполняет наружная мембрана клеточной стенки, через которую молекулы транспортируются только по механизму пассивной диффузии. Молекулы воды, некоторых газов (например O2, H2, N2) и углеводородов, концентрации которых во внешней среде выше, чем в клетке, проходят через ЦПМ внутрь клетки посредством пассивной диффузии. Движущей силой этого процесса служит градиент концентрации вещества по обе стороны мембраны. Основным соединением, поступающим в клетку и покидающим ее таким путем, является вода. Движение воды через мембрану, подчиняющееся законам пассивной диффузии, привело к выводу о существовании в мембране пор. Эти поры пока не удалось увидеть в электронный микроскоп, но некоторые данные о них были получены косвенными методами. Расчетным путем установлено, что поры должны быть очень мелкими и занимать небольшую часть поверхности ЦПМ. Высказывается предположение, что они не являются стабильными структурными образованиями, а возникают в результате временных перестроек молекулярной организации мембраны. Большинство (если не все) гидрофильных веществ поступает в клетку за счет функционирования систем, в состав которых входят специальные переносчики (транслоказы, или пермеазы), так как скорость физической диффузии этих веществ через гидрофобный слой мембраны очень невелика. Переносчики — вещества белковой природы, локализованные в мембране и характеризующиеся высокой субстратной специфичностью, — связываясь с субстратом, подвергаются конформационным изменениям и вследствие этого приобретают способность к перемещению субстрата с одной стороны ЦПМ на другую. Известен механизм транспорта, получивший название облегченной диффузии, требующий для переноса веществ через мембрану участия транслоказ. Перенос веществ в этом случае происходит по градиенту их концентрации и не требует энергетических затрат. Этот механизм транспорта не получил широкого распространения у прокариот. Основным механизмом избирательного переноса веществ через ЦПМ прокариот является активный транспорт, позволяющий " накачивать" в клетку молекулы и ионы против их концентрационных и электрических градиентов. Этот процесс, так же как и облегченная диффузия, протекает при участии локализованных в ЦПМ переносчиков белковой природы с высокой специфичностью к субстрату, но в отличие от облегченной диффузии для движения против электрохимического градиента требует затрат метаболической энергии. Транспорт такого рода должен быть, поэтому сопряжен с реакциями, продуцирующими энергию в химической или электрохимической форме. Во всех описанных выше путях переноса веществ через ЦПМ они поступают в клетку в химически неизмененном виде. У прокариот известны системы транспорта, с помощью которых осуществляется поступление в клетку ряда сахаров, при этом процесс их переноса через мембрану сопровождается химической модификацией молекул. Так происходит, например, поступление в клетки многих прокариот молекул глюкозы, в процессе которого они фосфорилируются. Цитозоль и рибосомы Содержимое клетки, окруженное ЦПМ, называется цитоплазмой. Фракция цитоплазмы, имеющая гомогенную консистенцию и содержащая набор растворимых РНК, ферментных белков, продуктов и субстратов метаболических реакций, получила название цитозоля. Другая часть цитоплазмы представлена разнообразными структурными элементами: внутрицитоплазматическими мембранами (если они есть), генетическим аппаратом, рибосомами и включениями разной химической природы и функционального назначения. Рибосомы — место синтеза белка — рибонуклеопротеиновые частицы размером 15 – 20 нм. Их количество в клетке зависит от интенсивности процессов белкового синтеза и колеблется от 5000 до 90 000. Общая масса рибосом может составлять примерно 1/4 клеточной массы, а количество рибосомальной РНК (рРНК) — 80 – 85 % всей бактериальной РНК. Отношение рРНК/белок в рибосомах Е. coli составляет 2: 1, у других прокариот оно может быть несколько сдвинуто в сторону преобладания белка. Рибосомы прокариот имеют константу седиментации 705, отчего получили название 70S-частиц. Они построены из двух неодинаковых субчастиц: 305- и 50S-субъединиц (Обозначения 30S, 50S, 70S — константы седиментации, характеризующие скорость, с которой эти частицы осаждаются в центрифуге при определенных стандартных условиях). 30S-частица содержит одну молекулу 16S-рРНК и в большинстве случаев по одной молекуле белка более 20 видов. 50S-субъединица состоит из двух молекул рРНК (235 и 55). В ее состав входят более 30 различных белков, также представленных, как правило, одной копией. Большая часть рибосомальных белков выполняет структурную функцию. Синтез белка осуществляется агрегатами, состоящими из рибосом, молекул информационной и транспортных РНК и называемыми полирибосомами, или полисомами. Последние могут находиться в цитоплазме или же быть связанными с мембранными структурами. Рост и способы размножения Под ростом прокариотной клетки понимают согласованное увеличение количества всех химических компонентов, из которых она построена. Рост является результатом множества скоординированных биосинтетических процессов, находящихся под строгим регуляторным контролем, и приводит к увеличению массы (а следовательно, и размеров) клетки. Но рост клетки не беспределен. После достижения определенных (критических) размеров клетка подвергается делению. Популярное:
|
Последнее изменение этой страницы: 2017-03-03; Просмотров: 1175; Нарушение авторского права страницы