|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Структура, химический состав и функции компонентов прокариотной клетки
Клетка прокариот обладает рядом принципиальных особенностей, касающихся как ее ультраструктурной, так и химической организации (рис. 2). Структуры, расположенные снаружи от ЦПМ (клеточная стенка, капсула, слизистый чехол, жгутики, ворсинки), называют обычно поверхностными структурами. Термином " клеточная оболочка" часто обозначают все слои, располагающиеся с внешней стороны от ЦПМ (клеточная стенка, капсула, слизистый чехол). ЦПМ вместе с цитоплазмой называется протопластом. Рассмотрим сначала строение, химический состав и функции поверхностных клеточных структур.
Рис. 2 Комбинированное изображение прокариотной клетки А — поверхностные клеточные структуры и внеклеточные образования: 1 — клеточная стенка; 2 — капсула; 3 — слизистые выделения; 4 — чехол; 5 — жгутики; 6 — ворсинки; Б — цитоплазматические клеточные структуры: 7 — ЦПМ; 8 — нуклеоид; 9 — рибосомы; 10 — цитоплазма; 11 — хроматофоры; 12 — хлоросомы; 13 — пластинчатые тилакоиды; 14 — фикобилисомы; 15 — трубчатые тилакоиды; 16 — мезосома; 17 — аэросомы (газовые вакуоли); 18 — ламеллярные структуры; В — запасные вещества: 19 — полисахаридные гранулы; 20 — гранулы поли-b-оксимасляной кислоты; 21 — гранулы полифосфата; 22 — цианофициновые гранулы; 23 — карбоксисомы (полиэдральные тела); 24 — включения серы; 25 — жировые капли; 26 — углеводородные гранулы (по Schlegel, 1972). Клеточная стенка Клеточная стенка — важный и обязательный структурный элемент подавляющего большинства прокариотных клеток, располагающийся под капсулой или слизистым чехлом или же непосредственно контактирующий с окружающей средой (у клеток, не содержащих этих слоев клеточной оболочки). На долю клеточной стенки приходится от 5 до 50 % сухих веществ клетки. Клеточная стенка служит механическим барьером между протопластом и внешней средой и придает клеткам определенную, присущую им форму. Концентрация солей в клетке, как правило, намного выше, чем в окружающей среде, и поэтому между ними существует большое различие в осмотическом давлении. Клеточная стенка чисто механически защищает клетку от проникновения в нее избытка воды. По строению и химическому составу клеточная стенка прокариот резко отличается от таковой эукариотных организмов. В ее состав входят специфические полимерные комплексы, которые не содержатся в других клеточных структурах. Химический состав и строение клеточной стенки постоянны для определенного вида и являются важным диагностическим признаком. В зависимости от строения клеточной стенки прокариоты, относящиеся к эубактериям, делятся на две большие группы. Было обнаружено, что если фиксированные клетки эубактерий обработать сначала кристаллическим фиолетовым, а затем йодом, образуется окрашенный комплекс. При последующей обработке спиртом в зависимости от строения клеточной стенки судьба комплекса различна. У так называемых грамположительных видов этот комплекс удерживается клеткой, и последние остаются окрашенными, у грамотрицательных видов, наоборот, окрашенный комплекс вымывается из клеток, и они обесцвечиваются. У некоторых эубактерий положительная реакция при окрашивании описанным выше способом свойственна только клеткам, находящимся в стадии активного роста. Выяснено, что окрашенный комплекс образуется на протопласте, но его удерживание клеткой или вымывание из нее при последующей обработке спиртом определяются особенностями строения клеточной стенки.
Рис. 3 Клеточная стенка грамположительных (А) и грамотрицательных (Б) эубактерий 1 — цитоплазматическая мембрана; 2 — пептидогликан; 3 — периплазматическое пространство; 4 — наружная мембрана: 5 — цитоплазма, в центре которой расположена ДНК. Клеточные стенки грамположительных и грамотрицательных эубактерий резко различаются как по химическому составу, так и по ультраструктуре (рис. 3). В состав клеточной стенки эубактерий входят семь различных групп химических веществ, при этом пептидогликан присутствует только в клеточной стенке. У грамположительных 1 эубактерий он составляет основную массу вещества клеточной стенки (от 40 до 90 %), у грамотрицательных — содержание пептидогликана значительно меньше (1 – 10 %). Клеточная стенка цианобактерий, сходная с таковой грамотрицательных эубактерий, содержит от 20 до 50 % этого гетерополимера. Этот способ был впервые предложен в 1884 г. датским ученым X. Грамом (Ch. Gram), занимавшимся окрашиванием тканей. Позднее он был использован для бактерий. Под электронным микроскопом клеточная стенка грамположительных эубактерий выглядит как гомогенный электронно-плотный слой, толщина которого колеблется для разных видов от 20 до 80 нм. У грамотрицательных эубактерий обнаружена многослойная клеточная стенка. Внутренний электронно-плотный слой толщиной порядка 2 – 3 нм состоит из пептидогликана. Снаружи к нему прилегает, как правило, волнистый слой (8 – 10 нм), имеющий характерное строение: две электронно-плотные полосы, разделенные электронно-прозрачным промежутком. Такой вид характерен для элементарных мембран. Поэтому трехконтурный внешний компонент клеточной стенки грамотрицательных эубактерий получил название наружной мембраны.
Рис. 4 Структура повторяющейся единицы пептидогликана клеточной стенки эубактерий 1, 2 — места полимеризации гликанового остова молекулы; 3 — место присоединения с помощью фосфодиэфирной связи молекулы тейхоевой кислоты в клеточной стенке грамположительных эубактерий; 4, 5 — места, по которым происходит связывание между гликановыми цепями с помощью пептидных связей; 6 — место ковалентного связывания (пептидная связь) с липопротеином наружной мембраны у грамотрицательных эубактерий; 7 — место действия лизоцима.
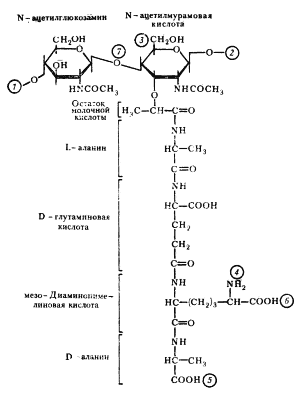
Клеточная стенка грамположительных эубактерий плотно прилегает к ЦПМ, в отличие от клеточной стенки грамотрицательных видов, компоненты которой (пептидогликановый слой и наружная мембрана) разделены электронно-прозрачным промежутком и четко отделены аналогичным образом от ЦПМ. Пространство между цитоплазматической: и наружной мембранами получило название периплазматического. Оно, как можно видеть из строения клеточных стенок обеих групп эубактерий, характерно только для грамотрицательных форм. Клеточная стенка грамположительных эубактерий. Основную массу клеточной стенки грамположительных эубактерий составляет специфический гетерополимер — пептидогликан. Полисахаридный остов молекулы построен из чередующихся остатков N-ацетилглюкозамина и N-ацетилмурамовой кислоты, соединенных между собой посредством b-1, 4-гликозидных связей (рис. 4). К N-ацетилмурамовой кислоте присоединен короткий пептидный хвост, состоящий из небольшого числа (обычно 4 – 5) аминокислот. У грамположительных эубактерий обнаружено более 100 различных химических типов пептидогликана. Большинство различий относится к пептидной части его молекулы. Две особенности пептидного хвоста заслуживают внимания: наличие аминокислот в D-форме (неприродная конфигурация) и высокое содержание аминокислот с двумя аминогруппами. Это имеет принципиальное значение для пространственной организации пептидогликана. Обе аминогруппы этих аминокислот могут участвовать в образовании пептидных связей, причем вторые аминогруппы — в формировании дополнительных пептидных связей между гетерополимерными цепочками.
Рис. 5 Пептидные мостики между гетерополимерными цепочками Г — N-ацетилглюкозамин; М — N-ацетилмурамовая кислота; ала — аланин; глу — глутаминовая кислота; лиз — лизин; ДАП — диаминопимелиновая кислота; гли — глицин; стрелками обозначено место действия пенициллина. В большинстве случаев в образовании пептидной связи участвует карбоксильная группа D-аланина одного тетрапептида и свободная аминогруппа диаминокислоты другого (рис. 5, А). Иногда связь между тетрапептидами разных гликановых цепей осуществляется с помощью других аминокислот (рис. 5, Б). Нетрудно себе представить, что этим способом можно " сшить" между собой множество гетерополимерных цепей. Частота " сшивок" различна, поскольку не все пептидные хвосты участвуют в формировании межцепочечных связей. Некоторые образуют ковалентные связи с другими химическими молекулами, входящими в состав клеточной стенки, и, наконец, часть тетрапептидных хвостов находится в свободном состоянии. Пептидогликан, окружающий протопласт грамположительных эубактерий, — это по существу одна гигантская молекула, " сшитая" с помощью гликозидных и пептидных связей. Именно последние обеспечивают ей трехмерную пространственную организацию. Кроме пептидогликана в состав клеточных стенок грамположительных эубактерий входит другой уникальный класс химических соединений — тейхоевые кислоты, представляющие собой полимеры, построенные на основе рибита (пятиатомного спирта) или глицерина (трехатомного спирта), остатки которых соединены между собой фосфодиэфирными связями (рис. 6). Некоторые свободные гидроксильные группы в молекулах спиртов могут быть замещены остатками D-аланина, глюкозы, N-ацетилглюкозамина и некоторых других сахаров. Тейхоевые кислоты ковалентно могут соединяться с N-ацетилмурамовой кислотой (см. рис. 4). Поскольку это длинные линейные молекулы, они могут пронизывать весь пептидогликановый слой, достигая внешней поверхности клеточной стенки. В этом случае, вероятно, они являются основными антигенами грамположительных эубактерий. Остающиеся свободные гидроксилы фосфорной кислоты придают тейхоевой кислоте свойства полианиона. Как полианионы тейхоевые кислоты определяют поверхностный заряд клетки. Сахарные компоненты тейхоевых кислот входят в состав рецепторов для некоторых бактериофагов и определяют возможность адсорбции фага на клеточной поверхности.
Рис. 6 Структурная формула глицеринтейхоевой кислоты Содержит чередующиеся остатки D-аланина и N-ацетилглюкозамина (по Rose, 1971). В составе клеточной стенки грамположительных эубактерий в небольших количествах также найдены полисахариды, белки и липиды. Для полисахаридов и липидов показана возможность ковалентного связывания с макромолекулами клеточной стенки, в отличие от белков, которые (у тех видов, где имеются) формируют на ее внешней поверхности отдельный слой. Таким образом, основными компонентами клеточной стенки грамположительных эубактерий являются три типа макромолекул: пептидогликаны, тейхоевые кислоты и полисахариды, которые с помощью ковалентных связей образуют сложную структуру с весьма упорядоченной пространственной организацией. Клеточная стенка бацилл, например Bacillus subtilis, приблизительно соответствует толщине 40 молекул пептидогликана. В целом клеточную стенку грамположительных эубактерий можно представить в виде губчатой структуры с порами диаметром примерно 1 – 6 нм. Возможность прохождения молекул через такую клеточную стенку определяется ее зарядом и размером пор. Клеточная стенка грамотрицательных эубактерий. У грамотрицательных эубактерий строение клеточной стенки намного сложнее, чем у грамположительных (см. рис. 3). В ее состав входит гораздо большее число макромолекул разного химического типа (см. табл. 1). Таблица 1/3 Химический состав клеточных стенок грамположительных и грамотрицательных эубактерий (по Rose, 1971; Freer, Salton, 1971)
бозначения: (–) — отсутствуют, (+) — присутствуют, (±) — присутствуют не у всех видов Пептидогликан образует только внутренний слой клеточной стенки, неплотно прилегая к ЦПМ. Для разных видов грамотрицательных эубактерий содержание этого гетерополимера колеблется в широких пределах. У большинства видов он образует одно- или двухслойную структуру, характеризующуюся весьма редкими поперечными связями между гетерополимерными цепями (рис. 7).
Рис. 7 Однослойная структура пептидогликана Линиями обозначены гетерополимерные цепочки, образованные чередующимися остатками N-ацетилглюкозамина (Г) и N-ацетилмурамовой кислоты (М), соединенными между собой b-1, 4-гликозидными связями. Кружочками обозначены аминокислоты пептидного хвоста. Химическая структура пептидогликана грамотрицательных эубактерий в основном сходна со структурой типичного пептидогликана грамположительных эубактерий (см. рис. 4; 5, А). Снаружи от пептидогликана располагается дополнительный слой клеточной стенки — наружная мембрана. Она состоит из фосфолипидов, типичных для элементарных мембран, белков, липопротеина и липополисахарида (рис. 8, А). Специфическим компонентом наружной мембраны является липополисахарид сложного молекулярного строения, занимающий около 30 – 40 % ее поверхности и локализованный во внешнем слое (рис. 8, 5).
Рис. 8 А. Клеточная стенка грамотрицательных эубактерий: 1 — цитоплазматическая мембрана; 2 — пептидогликановый слой; 3 — периплазматическое пространство; 4 — молекулы белков (заштрихована гидрофобная часть); 5 — фосфолипид; 6 — липополисахарид. Б. Строение молекулы липополисахарида: 1 — липид А; 2 — внутреннее полисахаридное ядро; 3 — наружное полисахаридное ядро; 4 — О-антиген. Белки наружной мембраны можно разделить на основные и минорные. Основные белки представлены небольшим числом различных видов, но составляют почти 80 % всех белков наружной мембраны. Одна из функций этих белков — формирование в мембране гидрофильных пор диаметром примерно 1 нм, через которые осуществляется неспецифическая диффузия молекул с массой до 600 – 900 Да7 (7Да — дальтон, или единица атомной массы, равен 1, 66033 x 10-27 кг). Это означает, что через такие поры могут проходить сахара, аминокислоты, небольшие олигосахариды и пептиды. Белки, пронизывающие наружную мембрану насквозь и образующие гидрофильные поры, называют поринами. Минорные белки наружной мембраны представлены гораздо большим числом видов. Их основная функция — транспортная и рецепторная. Примером минорных белков могут служить белки, ответственные за специфический транспорт в клетку железосодержащих соединений. Помимо слоев клеточной стенки, типичных для большинства грамотрицательных эубактерий, у некоторых представителей этой группы обнаружены дополнительные слои разной электронной плотности, располагающиеся с внешней стороны от наружной клеточной мембраны. Однако до настоящего времени не ясно, относятся ли они к клеточной стенке, являясь результатом ее последующего усложнения, или же представляют собой структурные элементы многослойного чехла. Популярное:
|
Последнее изменение этой страницы: 2017-03-03; Просмотров: 651; Нарушение авторского права страницы