
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Явление гетерозиса. Генетические механизмы гетерозиса.
Гетерозис — это свойство помесей и гибридов первого поколения (Fi) превосходить по биологическим и хозяйственно-полезным признакам исходные родительские формы. Впервые явление гетерозиса было описано И. Кёльрейтером, работавшим в Петербургской академии наук, на примере межвидового гибрида, который он получил в 1760 г., скрестив два разных вида махорки. Этот растительный гибрид оказался стерильным наподобие мула и автор назвал его «первым растительным мулом». Научный термин «гетерозис» появился гораздо позже. Его предложил американский исследователь Дж. Шелл в 1914 г. для обозначения мощности гибридов (эффекта скрещивания), и с тех пор он прочно вошел в научную литературу как синоним старого названия «гибридная сила». В настоящее время твердо установлено, что гетерозис проявляется не только при разведении животных и птиц, но и при селекции растений, а также микроорганизмов. Следовательно, гетерозис — явление общебиологическое. Каковы причины возникновения гетерозиса? На этот счет существует несколько точек зрения в виде отдельных самостоятельных гипотез. Одна из первых изложена в фундаментальном труде Чарльза Дарвина «Действие перекрестного опыления и самоопыления в растительном мире», в котором он изложил вопросы гетерозиса («гибридной силы»). Изучив опыт английских заводчиков по созданию новых пород сельскохозяйственных животных, Ч. Дарвин отметил, что родственное спаривание (инбридинг), которое применялось им для закрепления в потомстве желательных признаков выдающихся производителей, приводит, как и самоопыление у растений, к отрицательным последствиям — к депрессии, в то время как скрещивание повышает, как правило, жизнеспособность потомства за счет проявления эффекта скрещивания (гетерозиса). Дарвин предположил, что в основе этих двух явлений — инбредной депрессии и гетерозиса — лежит одна и та же причина — степень различия половых элементов, объединяющихся в процессе оплодотворения. Чем больше родительские формы, а следовательно, и их половые клетки различаются по своим биологическим особенностям, тем сильнее в потомстве проявляется гетерозис, и наоборот, отсутствие таких различий при близкородственном длительном спаривании приводит к нибредной депресии. Исходя из этих соображений, Ч. Дарвин провозгласил «великий закон природы», по которому, сточки зрения эволюции вида, скрещивание всегда полезно, а родственное спаривание (инбридинг — у животных, самоопыление — у растений) — вредно. Гипотеза доминирования (Джонс, 1972). В основе этой гипотезы лежит идея благоприятного действия доминантных генов — гетерозис проявляется в результате взаимодействия при скрещивании благоприятных доминантных факторов, имеющихся у исходных родительских форм. Предполагается, что при скрещивании происходит сочетание благоприятно действующих неаллельных доминантных генов и одновременное подавление ими действия различных вредных рецессивных аллелей, которые у разных линий, а тем более пород, находятся в разных локусах. При скрещивани доминантные аллели, внесенные одним родителем (линией), могут перекрывать рецессивные аллели, полученные гибридом от другой родительской формы (линии). Проявление гетерозиса возможно также и за счет явления эпистаза, когда отдельные неаллельные гены (эпистатический ген) подавляют не только «свои» рецессивные, но и «чужие» доминантные гены (гипостатический ген). Гипотеза доминирования общепризнана, однако, она не объясняет полностью все вопросы, возникающие в связи с проявлением гетерозиса. Так, если исходить из названной гипотезы, то теоретически следует ожидать, что при полигибридном скрещивании гетерозигота Аа будет в той или иной степени приближаться по продуктивности к гомозиготе АА — приближаться, но не превосходить ее. Однако в практике давно установлено, что гетерозигота может превосходить по мощности не только рецессивную родительскую форму, но и доминантную, то есть обоих своих родителей. Это явление получило в генетике даже специальное название — сверхдоминирование, или моногибридный гетерозис. Гипотеза гетерозиготности (сверхдоминирование). С точки зрения гипотезы гетерозиготности проявление гетерозиса объясняется, говоря современным языком, разнокачественностью членов одной и той же пары аллелей у гибридных организмов как результат скрещивания различающихся исходных родительских форм. Соединение при гибридизации разнокачественных гамет родителей уже само по себе стимулирует более быстрый рост гетерозиготных гибридов, их лучшее развитие и т. д. В результате гибрид по мощности превосходит исходные гомозиготные родительские формы как рецессивную, так и доминантную, что и обусловливает эффект сверхдоминирования. В то же время гомозиготность родителей оказывает угнетающее действие на жизнеспособность потомства, что выражается формулой Аа> АА> аа. Предполагается, что у гетерозиготы оба аллеля одного локуса выполняют различные функции, взаимно дополняя друг друга в биохимическом процессе. При этом эффект гетерозиса будет тем выше, чем в большей степени аллели каждого локуса различаются функционально между собой, чем больше они дополняют друг друга. Причины гетерозиса, указанные в этих гипотезах, возможно, действуют одновременно, однако их, по-видимому, не достаточно для всестороннего объяснения механизма возникновения гетерозиса как общебиологического явления. По этому поводу проф. М. В. Лобашов (1969) писал: «Трудно предположить, чтобы в основе столь сложного явления, как гетерозис, лежал единственно генетический механизм». Что же касается понимания самого механизма генного взаимодействия при гетерозисе, то по современным воззрениям различие между обеими гипотезами незначительно или его совсем нет. Гипотеза генетического баланса (академик ВАСХНИЛ Н. В. Турбин, 1961. С точки зрения этой гипотезы явление гетерозиса нельзя объяснить действием одной какой-либо генетической причины — это суммарный эффект. Гипотеза генетического баланса, принимая в обеих чертах отдельные положения ранее высказанных гипотез, больше внимания, однако, уделяет взаимовлиянию неаллельных генов, физическим и биохимическим факторам, а также внешней среде вообще, условиям выращивания гибридов в частности. Особое внимание придается цитоплазматическому влиянию. Предполагается, что плазматические различия между гаметами должны стимулировать жизненные процессы у гибридного организма. Сбалансированность систем генов делает популяции наиболее приспособительными и продуктивными в конкретных условиях среды. Следует отметить, что в последние годы все большее значение приобретает также биохимическая теория гетерозиса , согласно которой скрещивание приводит к увеличению гетерозиготности по мутациям, регулирующим синтез белка — отсюда проявление гетерозиса происходит за счет обогащения биохимических процессов в клетках и тканях гибридного организма. Значение изложенных выше гипотез неоспоримо, однако ни одна из них не может быть пока признанной в качестве общепринятой теории гетерозиса. Возможно, прав известный генетик Ф. Хатт, в своем высказывании: «Гетерозис все еще представляет одну из самых больших загадок генетики». Что же представляет собой гетерозис как генетическое явление? Каковы формы проявления гетерозиса? Гетерозис — это совокупность явлений, связанных с повышенной жизнеспособностью помесей (гибридов), которая, как установлено, проявляется уже на ранних стадиях развития (онтогенеза). У эмбрионов гибридных кур, например, еще в эмбриональном развитии усиливаются обменные процессы, ускоряется их развитие, в результате чего вывод и качество суточного молодняка бывают выше по сравнению с теми же показателями линейных кур (Злочевская, 1968). Гетерозис — явление неустойчивое (кратковременное), оно наиболее ярко (четко) проявляется лишь в первом поколении (F1) скрещивания. Помесные (гибридные) животные при их дальнейшем разведении не дают подобных себе гетерозисных потомков, онине остаются «константными» по гетерозису. Поэтому их не оставляют на племя, а реализуют на мясо. Следовательно, гетерозис нельзя закрепить наследственно, его нужно всякий раз получать заново. В отдельных случаях гетерозис можно поддерживать на относительно высоком уровне и в последующих поколениях, но в таких случаях используются специальные методы — переменное скрещивание и др. Считается, что угасание гетерозиса в последующих поколениях гибридов — результат рекомбинационных утерь. Формы проявления гетерозиса бывают различными. Обычно при скрещивании двух пород (А и В) уровень продуктивности помесного (АВ) потомства равен средней показателей продуктивности исходных пород Нередко продуктивность помесных (F1) животных оказывается значительно выше средней продуктивности родителей, а иногда она превышает показатели лучшей из родительских форм — абсолютный (истинный) гетерозис. В других случаях, однако, продуктивность помесей превышает показатели лишь одного из родителей, худшего — относительный гетерозис: где: Пг — признак гибрида; Пл — признак лучшей породы; Пм — признак материнской породы; По — признак отцовской породы. Конечно, из чисто практических соображений эффект гетерозиса представляет наибольший интерес лишь в том случае, когда гибридное потомство превышает по своей общей хозяйственно полезной ценности лучшего из родителей. Только в таких случаях скрещивание имеет экономический смысл. Поэтому селекционеры-практики под гетерозисом понимают свойство гибридов (F1) превосходить по определенным признакам лучшую из родительских форм. Некоторые ученые, учитывая специфику форм проявления гетерозиса, выделяют самостоятельные его типы: репродуктивный гетерозис — более высокая общая продуктивность животных, связанная с повышением плодовитости (фертильности) и более мощным развитием их репродуктивных органов; соматический гетерозис — более сильное развитие вегетативных частей (у растений), органов и частей тела (у животных); адаптивный гетерозис — повышенная жизнеспособность животных, их лучшая приспособляемость. У мулов, например, сильно выражен соматический гетерозис, то есть большая живая масса; выше тяговое усилие; повышенное долголетие; особая выносливость; но в то же время репродуктивная система недоразвита. Как правило, они бесплодны. Сказанное выше — это пример частного гетерозиса (мощное развитие касается не всего организма животного, а лишь его отдельных признаков) в отличие от общего гетерозиса, когда имеет место развитие общей массы тела животного, повышение метаболических процессов в организме в целом, что обеспечивает повышение его продуктивности (Красота, 1979). Следует отметить, что гетерозис проявляется у помесей и гибридов — межвидовых, межпородных, межлинейных — по ограниченному числу признаков. Он никогда не проявляется по сумме всех родительских признаков. Помеси (гибриды) превосходят своих родителей не по всем показателям продуктивности, не по всем признакам, не по их сумме, а лишь частично, по отдельным признакам (или группе признаков) или даже по отдельно взятому признаку. Метисные куры первого поколения, полученные от скрещивания петухов мясных пород с курами яйценоских (легких) пород, могут превосходить исходные родительские формы по яйценоскости, но по живой массе они занимают промежуточное положение. Отсюда под гетерозисом следует понимать превосходство потомства — помесей или гибридов — над родительскими формами не по всем, а лишь по определенным, конкретным признакам.
Литература 1. Вавилов Н. И. Центры происхождения культурных растений.— Тр. по прикладной ботанике и селекции, 1926, т. XVI, с. 5—138. 2. Гершензон С. М. Основы современной генетики.— Киев, 1979.— 506 с. 3. Лобашев М. Е., Ватти К. В., Тихомирова М. М. Генетика с основами селекции.— М., 1979.— 304 с. 4. Рокицкий П. Ф. Некоторые этапы развития генетики животных в СССР и ее связи с селекцией.— В сб.: Генетические основы селекции животных. М., 1969, с. 9—25. ГЕНЕТИЧЕСКИЕ ОСНОВЫ СЕЛЕКЦИИ (4 часа) Лекция 20 Методы отбора, их значение Цель лекции: ознакомить учащихся с видами отбора, с успехами отечественных селекционеров в создании сортов растений и пород животных, с использованием достижений генетики в повышенной эффективности селекционного процесса. План лекции: 1. Методы отбора, их значение 2. Индивидуальный и массовый отборы. 3. Подбор
Методы отбора, их значение Отбор и подбор — основные приемы качественного улучшен ия животных. Отбор предусматривает выделение особей, обладающих желательными свойствами и признаками, которых оставляют для дальнейшего разведения. Животных, не отвечающих требованиям отбора, используют для производства продукции или выбраковывают. Это способствует улучшению из поколения в поколение желательных качеств животных. Однако для достижения успехов в улучшении их качеств одного отбора недостаточно. Весьма важное значение имеет эффективное использование лучших из отобранных животных путем их подбора для спаривания. Только при сочетании систематического отбора с целенаправленным подбором может быть достигнуто существенное повышение продуктивных и племенных качеств животных. Ведущая роль отбора в создании новых форм животных и растений обстоятельно разработана Ч. Дарвином. Им выделены отбор естественный и искусственный. Естественный отбор. Происходит он под воздействием естественных условий существования и направлен на лучшую выживаемость тех или иных форм. При естественном отборе получают преимущество формы, у которых в силу изменчивости появились новые признаки и свойства, благоприятствующие их выживанию в конкретных природных условиях. Наибольшее действие на животных естественный отбор оказывал на первых этапах одомашнения, а также при содержании их в условиях, близких к природным. При содержании сельскохозяйственных животных в современных помещениях с регулируемым микроклиматом и обеспечении их кормами влияние естественного отбора значительно ослабляется. Искусственный отбор. В отличие от естественного искусственный отбор осуществляется человеком и направлен на улучшение продуктивных и племенных качеств животных. Поэтому искусственный отбор не всегда согласуется с естественным. В частности, искусственный отбор на высокую продуктивность находится иногда в противоречии с естественным отбором на выживаемость животных в природных условиях. На первых этапах одомашнения животных человек, отбирая лучших особей, не ставил какой-либо цели по выведению животных, обладающих определенными качествами. Такой отбор — бессознательный. В последующем искусственный отбор получил направленный характер и преследовал цель выведения животных, отличающихся желательными качествами. Он назван Ч. Дарвином методическим. В настоящее время методический отбор приразведении животных получил почти повсеместное распространение. Действие отбора обусловлено наследственностью животных. Следует, однако, иметь в виду, что отклонения родителей по развитию признака от среднего значения для конкретной популяции (стадо, отара, завод и др.) наследуются потомками не в полной мере. У потомства наблюдается возврат к средней величине по стаду. Это явление называют регрессией. От лучших родителей рождаются, как правило, лучшие потомки, но их преимущество по развитию признака над средним его значением для популяции выражено в меньшей степени, чем преимущество родителей. В то же время потомство худших животных превосходит по развитию признака своих родителей. Явление регрессии обусловлено влиянием на развитие признака у потомков не только их родителей, но и более далеких предков, а также зависит от многообразия комбинаций и взаимодействия генов при половом размножении. Эффективность отбора определяется наследственной обусловленностью признаков и их изменчивостью, числом признаков, по которым он ведется, и точностью их измерения, интенсивностью отбора, численностью популяции, условиями кормления и содержания животных. В стаде или другой популяции животных, характеризующейся большей однородностью по развитию селекционируемых признаков, отбор будет менее эффективным, чем в стаде с высокой изменчивостью признака, свидетельствующей о разнообразии составляющих его животных. При увеличении числа признаков, по которым проводят отбор, эффективность его по развитию каждого признака снижается. По количественным (полигенным) признакам это снижение эффективности отбора пропорционально
где n — число признаков. Так, при увеличении числа признаков с 1 до 4 эффективность отбора по каждому признаку будет в 2 раза меньше
Следует, что наиболее высокий суммарный эффект по количеству молочного жира в годовом удое был получен при селекции коров по удою и значительно меньше при селекции по удою и содержанию жира в молоке, а также по жирности молока. При проведении отбора по нескольким признакам необходимо учитывать их взаимосвязь, характер их корреляции. При положительной корреляции отбор по одному признаку приводит к улучшению другого, взаимосвязанного с ним признака. Например, селекция по жирномолочности приводит часто к повышению содержания белка в молоке. При отрицательной корреляции между признаками селекция по обоим признакам возможна, но сопряжена с большими трудностями. При одновременном совершенствовании стада по таким признакам необходимо широко использовать животных, у которых оба признака хорошо выражены. Следует иметь в виду, что односторонний отбор по развитию какого-либо одного, хотя и очень ценного, признака может привести к ухудшению других. Так, известный овцевод П. Д. Мазаев проводил селекцию тонкорунных овец только по длине шерсти, недостаточно учитывая развитие других признаков. По селекционируемому признаку он достиг значительных успехов, но при этом наступило резкое ослабление конституции животных. Они стали более мелкими, ухудшился их экстерьер и снизилась жизнеспособность. При селекции животных по любому признаку надо обязательно учитывать и их конституцию, не допуская в процессе селекции ее ослабления. Число селекционируемых признаков ограничивают наиболее хозяйственно важными, предъявляя к развитию других признаков минимальные требования, чтобы не допустить их ухудшения. Большая численность животных в стаде или другой популяции способствует повышению эффективности отбора, так как позволяет выделять животных желательного типа из большего их числа. В таком же направлении действует интенсивность отбора. Чем она выше и чем больше по развитию признака отличаются животные отобранной группы от средних показателей по стаду (эта разница называется селекционным дифференциалом), тем больше оснований получить В потомстве особей с повышенной продуктивностью. Большое влияние на эффективность отбора по количественным признакам оказывают условия кормления и содержания животных. Только в тех случаях, когда эти условий благоприятствуют развитию селекционируемого признака, отбор будет эффективны. В частности, отбор коров по молочной продуктивности не будет эффективным при недостаточном и неполноценном их кормлении. При неудовлетворительных условиях кормления и содержания продуктивность всех животных будет невысокой. В зоотехнической практике оценку и отбор животных проводят по фенотипу и генотипу, учитывая происхождение, индивидуальные свойства и качество их потомства. Оценку по происхождению проводят по фенотипу и генотипу предков и ближайших родственников. По происхождению можно определить породность животного, его принадлежность к той или иной генетической группе (линия, семейство) и методы подбора, применявшиеся при его получении. При оценке по происхождению учитывают продуктивность предков и боковых родственников (полных сестер и братьев — сибсов, а также полусестер и полубратьев — полусибсов). Учет продуктивности сибсов и полусибсов дает представление о генотипе прямых предков животных. Происхождение животных устанавливают по родословной, в которой указаны сведения о матерях и отцах животных, их дедах и бабках, прадедах и прабабках и о более отдаленных предках. В родословной предков животного указывают в соответствующих рядах, которые обозначают римскими цифрами I, II, III и т. д. В I ряд родословной записывают родителей, во II — дедов и бабок, в III — прадедов и прабабок и т.д. Первая ступень отбора — оценка животного по происхождению. Она преследует цель определить дальнейшее назначение животного — выращивание на племя или на мясо и т. д. Вторая ступень отбора — оценка животного по индивидуальным качествам: по развитию и в последующем по его продуктивности. Учитывают в первую очередь те признаки, по которым проводят селекцию: энергию роста, уровень продуктивности, телосложение, плодовитость. Эти данные характеризуют фенотип животного. Третья ступень отбора — оценка племенных достоинств животного, его генотипа по качеству полученного от него потомства. Такую оценку проводят по массовым данным зоотехнического учета и результатам испытания животных по продуктивности потомства. Оценка животных по качеству потомства позволяет более достоверно судить об их племенной ценности. При оценке животных по качеству потомства применяют несколько приемов сравнения потомков по продуктивности — с матерями, со сверстницами, со стандартом по породе, со средними показателями по стаду. Сравнение продуктивности потомков и матерей применимо в хозяйствах, где условия кормления и содержания животных в течение ряда поколений обеспечивают развитие селекционируемого признака и мало изменяются. Сравнение потомков животного со сверстницами или средними показателями по стаду имеет большое значение для решения вопроса о дальнейшем использовании животного в конкретном стаде или группе стад с близким уровнем продуктивности и имеющих сходную генеалогическую структуру. В племенных хозяйствах и на племенных фермах результаты отбора ежегодно получают отражение в бонитировке животных. Последняя предусматривает оценку животных по развитию ведущих признаков. Признаки, которые учитываются при бонитировке животных, определяются их видовыми, породными особенностями и направлением селекции. По результатам бонитировки животных распределяют на классы. Для крупного рогатого скота приняты следующие классы: элита-рекорд, элита, I и II классы; для свиней и лошадей — элита, I и II классы; для овец —I, II и III классы. На основании данных бонитировки определяют дальнейшее назначение животного и составляют план спариваний. Популярное:
|
Последнее изменение этой страницы: 2017-03-08; Просмотров: 1399; Нарушение авторского права страницы
 В таких случаях говорят о гипотетическом (вероятном) гетерозисе.
В таких случаях говорят о гипотетическом (вероятном) гетерозисе. 

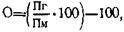
 ,
, 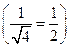 .
.