|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Происхождение жизни: гипотеза панспермии и абиогенного происхождения жизни. Главные этапы возникновения и развития жизни.Стр 1 из 22Следующая ⇒
Существуют две главные гипотезы, по-разному объясняющие появление жизни на Земле. Согласно гипотезе панспермии, жизнь занесена из космоса либо в виде спор микроорганизмов, либо путем намеренного «заселения» планеты разумными пришельцами из других миров. Прямых свидетельств в пользу космического происхождения жизни нет. Космос, однако, наряду с вулканами мог быть источником низкомолекулярных органических соединений, раствор которых послужил средой для развития жизни. Современной наукой возраст Земли оценивается в 4, 5—4, 6 млрд. лет. Появление на планете первых водоемов, с которыми связывают зарождение жизни, отстоит от настоящего времени на 3, 8—4 млрд. лет. Полагают, что около 3, 8 млрд. лет назад жизнь могла стать определяющим фактором планетарного круговорота углерода. В породах вблизи местечка Фиг-Три (Южная Африка), имеющих возраст более 3, 5 млрд. лет, обнаружены бесспорные следы жизнедеятельности микроорганизмов. Таким образом, процесс образования примитивных живых существ шел относительно быстро. Ускорению процесса могло способствовать то, что простейшие органические вещества были из нескольких источников: абиогенно образующиеся в первичной атмосфере и в то же время поступающие с оседающей на поверхность планеты космической и вулканической пылью. Подсчитано, что Земля, проходя через пылевое облако в течение 1 млрд. лет, могла получить с космической пылью 10 млрд. т органического материала. Это всего в 300 раз меньше суммарной биомассы современных наземных организмов (3 • 1012 т). Вулкан за одно извержение выбрасывает до 1000 т органических веществ. Согласно второй гипотезе, жизнь возникла на Земле, когда сложилась благоприятная совокупность физических и химических условий, сделавших возможным абиогенное образование органических веществ из неорганических. В середине прошлого столетия Л. Пастер окончательно доказал невозможность самозарождения жизни в теперешних условиях. В 20-х годах текущего столетия биохимики А. И. Опарин и Дж. Холдейн предположили, что в условиях, имевших место на планете несколько миллиардов лет назад, образование живого вещества было возможно. К таким условиям они относили наличие атмосферы восстановительного типа, воды, источников энергии (в виде ультрафиолетового (УФ) и космического излучения, теплоты остывающей земной коры, вулканической деятельности, атмосферных электрических явлений, радиоактивного распада), приемлемой температуры, а также отсутствие других живых существ. Главные этапы на пути возникновения и развития жизни, по-видимому, состоят в: 1) образовании атмосферы из газов, которые могли бы служить «сырьем» для синтеза органических веществ (метана, оксида и диоксида углерода, аммиака, сероводорода, цианистых соединений), и паров воды; 2) абиогенном (т.е. происходящем без участия организмов) образовании простых органических веществ, в том числе мономеров биологических полимеров — аминокислот, Сахаров, азотистых оснований, АТФ и других мононуклеотидов; 3) полимеризации мономеров в биологические полимеры, прежде всего белки (полипептиды) и нуклеиновые кислоты (полинуклеотиды); 4) образовании предбиологических форм сложного химического состава — протобионтов, имеющих некоторые свойства живых существ; 5) возникновении простейших живых форм, имеющих всю совокупность главных свойств жизни, —примитивных клеток; 6) биологической эволюции возникших живых существ. Возможность абиогенного образования органических веществ, включая мономеры биологических полимеров, в условиях, бывших на Земле около 4 млрд. лет назад, доказана опытами химиков. В лабораторных условиях при пропускании электрических разрядов через различные газовые смеси, напоминающие примитивную атмосферу планеты, а также при использовании других источников энергии ученые получали среди продуктов реакций аминокислоты (аланин, глицин, аспарагиновую кислоту), янтарную, уксусную, молочную кислоты, мочевину, азотистые основания (аденин, гуанин), АДФ и АТФ. Низкомолекулярные органические соединения накапливались в водах первичного океана в виде первичного бульона или же адсорбировались на Поверхности глинистых отложений. Последнее повышало концентрацию этих веществ, создавая тем самым лучшие условия для полимеризации. Возможность полимеризации низкомолекулярных соединений с образованием полипептидов и полинуклеотидов (определяющая следующий этап на пути возникновения жизни) непосредственно в первичном бульоне вызывает сомнения по термодинамическим соображениям. Водная среда благоприятствует реакции деполимеризации. Ученые предполагают, что образование полипептидов и полинуклеотидов могло происходить в пленке из низкомолекулярных органических соединений в безводной среде, например на склонах вулканических конусов, покрытых остывающей лавой. Это предположение находит подтверждение в опытах. Выдерживание в течение определенного времени при 130°С сухой смеси аминокислот в сосудах из кусков лавы приводило к образованию полипептидов. Образующиеся описанным образом биополимеры смывались ливневыми потоками в первичный бульон, что защищало их от разрушающего действия УФ-излучения, которое в то время из-за отсутствия в атмосфере планеты озонового слоя было очень жестким. По мере повышения концентрации полипептидов, полинуклеотидов и других органических соединений в первичном бульоне сложились условия для следующего этапа — самопроизвольного возникновения предбиологических форм сложного химического состава, или протобионтов. Предположительно они могли быть представлены коацерватами (А. И. Опарин) или микросферами (С. Фоке). Это коллоидные капли с уплотненным поверхностным слоем, имитирующим мембрану, содержимое которых составляли один или несколько видов биополимеров. Возможность образования в коллоидных растворах структур типа коацерватов или микросфер доказана опытным путем. При определенных условиях коацерваты проявляют некоторые общие свойства живых форм. Они способны до известной степени избирательно поглощать вещества из окружающего раствора. Часть продуктов химических реакций, проходящих в коацерватах с участием поглощаемых веществ, выделяется ими обратно в среду. Происходит процесс, напоминающий обмен веществ. Накапливая вещества, коацерваты увеличивают свой объем (рост). По достижении определенных размеров они распадаются на части, сохраняя при этом некоторые черты исходной химической организации (размножение). Поскольку устойчивость коацерватов различного химического состава различна, среди них происходит отбор. Перечисленные выше свойства ученые усматривают у протобионтов. Протобионты представляются как обособленные от окружающей среды, открытые макромолекулярные системы, возникавшие в первичном бульоне и способные к примитивным формам роста, размножения, обмена веществ и предбиологическому химическому отбору. Предбиологическая эволюция протобионтов осуществлялась в трех главных направлениях. Важное значение имело совершенствование каталитической (ферментной) функции белков. Один из путей, дающих требуемый результат, заключается, по-видимому, в образовании комплексов металлов с органическими молекулами. Так, включение железа в порфириновое кольцо Гемоглобина увеличивает его каталитическую активность в сравнении с активностью самого железа в растворе в 1000 раз. Развивалось такое свойство биологического катализа, как специфичность. Во-вторых, исключительная роль в эволюции протобионтов принадлежит приобретению полинуклеотидами способности к самовоспроизведению, что сделало возможным передачу информации от поколения к поколению, т.е. сохранение ее во времени. В основе этой способности лежит матричный синтез. Механизм матричного синтеза был использован также для переноса информации с полинуклеотидов на полипептиды. Третье главное направление эволюции протобионтов состояло в возникновении мембран. Отграничение от окружающей среды мембраной с избирательной проницаемостью превращает протобионт в устойчивый набор макромолекул, стабилизирует важные параметры обмена веществ на основе специфического катализа. Разделение функций хранения и пространственно-временной передачи информации, с одной стороны (нуклеиновые кислоты), и использование ее для организации специфических структуры и обмена веществ — с другой (белки); появление молекулярного механизма матричного синтеза биополимеров; освоение эффективных систем энергообеспечения жизнедеятельности (АТФ); образование типичной биологической мембраны — все это привело к возникновению живых существ, которые поначалу были представлены примитивными клетками. С момента появления клеток предбиологический химический отбор уступил место биологическому отбору. Дальнейшее развитие жизни шло согласно законам биологической эволюции. Переломным моментом на этом пути было возникновение клеток эукариотического типа, многоклеточных организмов, человека. Наряду с рассмотренными выше, предлагались и другие гипотезы происхождения жизни (см. 3.6.4.1), которые здесь в деталях не рассматриваются. 5.Типы клеточной организации. Строение про- и эукариотических клеток.
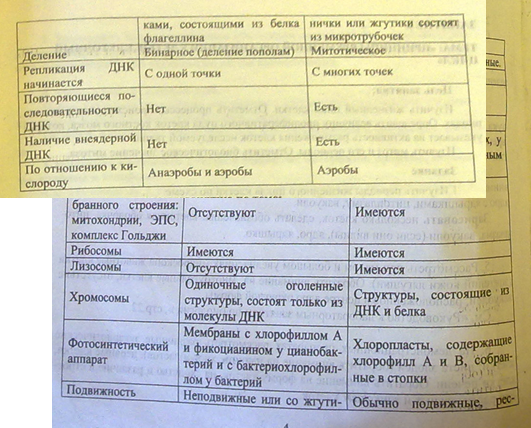
В настоящее время установлены 2 вида клеточной организации: прокариоты и эукариоты. Они существенно отличаются друг от друга. К прокариотическим организмам относят бактерии, СЗО и архебактерии (бактерии, выживающие в крайне тяжелых условиях). 0, 5-0, 3 мкм – размер клетки (см. таблицу в альбоме). Генетическая информация в одной хромосоме – двуцепочечная ДНК, кольцевой формы. Состав хромосом: нет гистоновых белков. Хромосома «голая». Распространены повсеместно. Короткая регенерация, короткое время размножения, быстрый рост, большое биохимическое разнообразие. Эукариотические клетки имеют сильно разветвленные внутриклеточные мембраны. Ядра содержат ядрышки и хромосомы (количество хромосом больше 2). В состав хромосом также входят белки-гистоны, РНК и др. эукариотические клетки способны существовать вместе с другими эукариотическими клетками и являются субъединицами многоклеточного организма. Прокариоты и эукариоты относятся к кислороду по-разному. Большинство прокариот – облигатные анаэробы, реже – факультативные анаэробы, есть и облигатные аэробы. Среди эукариот – единообразие – облигатные аэробы. Прокариоты возникли в период, когда содержание в среде кислорода изменялось, к моменту возникновения эукариот количество его было высоким и стабильным. Между прокариотами и эукариотами прочные эволюционные связи. У них сходные метаболические пути. У прокариот – брожение, у эукариот – гликолиз. Реакции похожи, механизм почти один и тот же. Анаэробное брожение как источник энергии возникло на ранних стадиях эволюции. С появлением кислорода появилась возможность более эффективного процесса окисления – 36 молекул АТФ из 1 молекулы глюкозы – окислительное фосфорилирование. Причем у эукариот оба процесса имеют место. Поэтому КПД – 38АТФ. Наличие обоих процессов имеет большое значение, один процесс может временно компенсировать другой. СЗО осуществляют аэробный фотосинтез. Предполагают, что цианобактерии способствовали накоплению кислорода в первичной атмосфере (около 1, 5 млрд. лет назад). 6.Гипотезы происхождения эукариотических клеток (симбиотическая, инвагинационная).
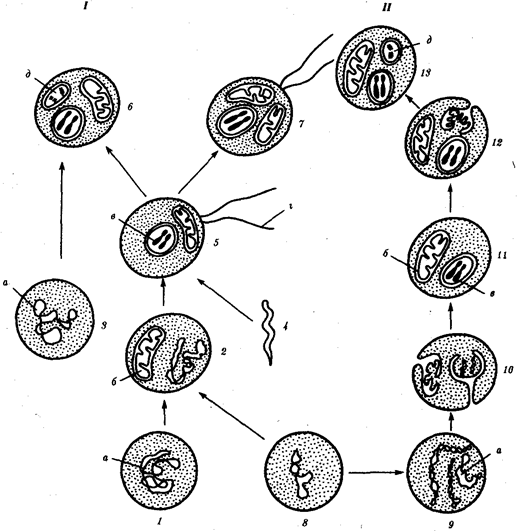
Ископаемые останки клеток эукариотического типа обнаружены в породах, возраст которых не превышает 1, 0—1, 4 млрд. лет. Более позднее возникновение, а также сходство в общих чертах их основных биохимических процессов (самоудвоение ДНК, синтез белка на рибосомах) заставляют думать о том, что эукариотические клетки произошли от предка, имевшего прокариотическое строение. Наиболее популярна в настоящее время симбиотическая гипотеза происхождения эукариотических клеток, согласно которой (рис. 1.4) основой, или клеткой-хозяином, в эволюции клетки эукариотического типа послужил анаэробный прокариот, способный лишь к амебоидному движению. Переход к аэробному дыханию связан с наличием в клетке митохондрии, которые произошли путем изменений симбионтов — аэробных бактерий, проникших в клетку-хозяина и сосуществовавших с ней.
Рис. 1.4. Происхождение эукариотической клетки согласно симбиотической (I) и инвагинационной (II) гипотезам: 1 — анаэробный прокариот (клетка-хозяин), 2 — прокариоты, имеющие митохондрии, 3 — сине-зеленая водоросль (презумптивный хлоропласт), 4 —сиирохетообразная бактерия (презумпгивный жгутик), 5 — примитивный эукариот со жгутиком, 6 — растительная клетка, 7 — животная клетка со жгутиком, 8 — аэробный прокариот (презумптивная митохондрия), 9 — аэробный прокариот (клетка-родоначальница согласно гипотезе II), 10 —инвагинации клеточной оболочки, давшие ядро и митохондрии, 11 — примитивный эукариот 12 — впячивание клеточной оболочки, давшее хлоропласт, 13 — растительная клетка; а—ДНК прокариотической клетки, б — митохондрия, в —ядро эукариотической клетки, г —жгутик, д — хлоропласт Сходное происхождение предполагают для жгутиков, предками которых служили симбионты-бактерии, имевшие жгутик и напоминавшие современных спирохет. Приобретение клеткой жгутиков имело наряду с освоением активного способа движения важное следствие общего порядка. Предполагают, что базальные тельца, которыми снабжены жгутики, могли эволюционировать в центриоли в процессе возникновения механизма митоза. Способность зеленых растений к фотосинтезу обусловлена присутствием в их клетках хлоропластов. Сторонники симбиотической гипотезы считают, что симбионтами клетки-хозяина, давшими начало хлоропластам, послужили прокариотические синезеленые водоросли. Серьезным доводом в пользу симбиотического происхождения митохондрий, центриолей и хлоропластов является то, что перечисленные органеллы имеют собственную ДНК. Вместе с тем белки бациллин и тубулин, из которых состоят жгутики и реснички соответственно современных прокариот и эукариот, имеют различное строение. У бактерий не найдено также структур со свойственной жгутикам, ресничкам, базальным тельцам и центриолям эукариотических клеток комбинацией микротрубочек: «9 + 2» или «9 + 0». Внутриклеточные мембраны гладкой и шероховатой цитоплазматической сети, пластинчатого комплекса, пузырьков и вакуолей рассматривают как производные наружной мембраны ядерной оболочки, которая способна образовывать впячивания. Центральным и трудным для ответа является вопрос о происхождении ядра. Предполагают, что оно также могло образоваться из симбионта-прокариота. Увеличение количества ядерной ДНК, во много раз превышающее в современной эукариотической клетке ее количество в митохондрий или хлоропласте, происходило, по-видимому, постепенно путем перемещения групп генов из геномов симбионтов. Нельзя исключить, однако, что ядерный геном формировался путем наращивания генома клетки-хозяина (без участия симбионтов). Согласно инвагинационной гипотезе, предковой формой эукариотической клетки был аэробный прокариот (рис. 1.4). Внутри такой клетки-хозяина находилось одновременно несколько геномов, первоначально прикреплявшихся к клеточной оболочке. Органеллы, имеющие ДНК, а также ядро, возникли путем впячивания и отшнуровывания участков оболочки с последующей функциональной специализацией в ядро, митохондрий, хлоропласты. В процессе дальнейшей эволюции произошло усложнение ядерного генома, появилась система цитоплазматических мембран. Инвагинационная гипотеза хорошо объясняет наличие в оболочках ядра, митохондрий, хлоропластов, двух мембран. Однако она не может ответить на вопрос, почему биосинтез белка в хлоропластах и митохондриях в деталях соответствует таковому в современных прокариотических клетках, но отличается от биосинтеза белка в цитоплазме эукариотической клетки. История показала, что эволюционные возможности клеток эукариотического типа несравнимо выше, чем прокариотического. Ведущая роль здесь принадлежит ядерному геному эукариот, который во много раз превосходит по размерам геном прокариот. Количество генов у бактерии и в клетке человека, например, соотносится как 1: (100-1000). Важные отличия заключаются в диплоидности эукариотических клеток благодаря наличию в ядрах двух комплектов генов, а также в многократном повторении некоторых генов. Это расширяет масштабы мутационной изменчивости без угрозы резкого снижения жизнеспособности, эволюционно значимым следствием чего является образование резерва наследственной изменчивости. При переходе к эукариотическому типу усложняется механизм регуляции жизнедеятельности клетки, что на уровне генетического материала проявилось в увеличении относительного количества регуляторных генов, замене кольцевых «голых» молекул ДНК прокариот хромосомами, в которых ДНК соединена с белками. В итоге стало возможным считывать биологическую информацию по частям с разных групп генов в разном их сочетании в различных типах клеток и в разное время. В бактериальной клетке, напротив, одновременно считывается до 80—100% информации генома. В клетках взрослого человека в разных его органах транскрибируется от 8—10% (печень, почка) до 44% (головной мозг) информации. Использованию биологической информации частями принадлежит исключительная роль в эволюции многоклеточных организмов, так как именно это позволяет разным группам клеток специализироваться по различным функциональным направлениям. Большое значение при переходе к многоклеточности имело наличие у эукариотических клеток эластичной оболочки, что необходимо для образования устойчивых клеточных комплексов. Среди цитофизиологических особенностей эукариот, увеличивающих их эволюционные возможности, необходимо назвать аэробное дыхание, которое также послужило предпосылкой для развития многоклеточных форм. Интересно, что сами эукариотические клетки появились на Земле после того, как концентрация O2 в атмосфере достигла 1% (точка Пастера). Названная концентрация является необходимым условием аэробного дыхания. В условиях усложнения генетического аппарата эукариот, увеличения суммарного количества ДНК и распределения ее по хромосомам трудно переоценить значение возникновения в эволюции митоза как механизма воспроизведения в поколениях генетически сходных клеток. Появление вследствие эволюционных преобразований митоза такого способа деления клеток, как мейоз, дающего возможность сохранить постоянство хромосом в ряду поколений, наилучшим образом решило проблему размножения многоклеточных организмов. Связанный с мейозом переход к половому размножению усилил эволюционную роль комбинативной изменчивости, способствовал увеличению скорости эволюции. Благодаря отмеченным особенностям за 1 млрд. лет эволюции эукариотический тип клеточной организации дал широкое разнообразие живых форм от одноклеточных простейших до млекопитающих и человека.
7.Иерархические уровни организации жизни. Проявления главных свойств жизни на различных уровнях её организации.
УРОВНИ ОРГАНИЗАЦИИ ЖИЗНИ
Живая природа является целостной, но неоднородной системой, которой свойственна иерархическая организация. Под системой, в науке понимают единство, или целостность, составленное из множества элементов, которые находятся в закономерных отношениях и связях друг с другом. Главные биологические категории, такие, как геном (генотип), клетка, организм, популяция, биогеоценоз, биосфера, представляют собой системы. Иерархической называется система, в которой части, или элементы, расположены в порядке от низшего к высшему. Так, в живой природе биосфера слагается из биогеоценозов, представленных популяциями организмов разных видов, а тела организмов имеют клеточное строение. Иерархический принцип организации позволяет выделить в живой природе отдельные уровни, что удобно с точки зрения изучения жизни как сложного природного явления. В медико-биологической науке широко используют классификацию уровней в соответствии с важнейшими частями, структурами и компонентами организма, являющимися для исследователей разных специальностей непосредственными объектами изучения. Такими объектами могут быть организм как таковой, органы, ткани, клетки, внутриклеточные структуры, молекулы. Выделение уровней рассматриваемой классификации хорошо согласуется с разрешающей способностью методов, которыми пользуются биологи и врачи: изучение объекта невооруженным глазом, с помощью лупы, светооптического микроскопа, электронного микроскопа, современных физико-химических методов. Очевидна связь этих уровней и с типичными размерами изучаемых биологических объектов (табл. 1.1).
Таблица 1.1. Уровня организации (изучения), выделяемые в многоклеточном организме (по Э. Дс. Робертсу и др., 1967, с изменениями)
Взаимопроникновение идей и методов различных областей естествознания (физики, химии, биологии), возникновение наук на стыке этих областей (биофизика, биохимия, молекулярная биология) повлекли за собой расширение классификации, вплоть до выделения молекулярного и электронно-атомного уровней. Медико-биологические исследования, проводимые на этих уровнях, уже сейчас дают практический выход в здравоохранение. Так, приборы, основанные на явлениях электронного парамагнитного и ядерного магнитного резонанса, с успехом применяют для диагностики заболеваний и состояний организма. Возможность исследовать фундаментальные биологические процессы, происходящие в организме, на клеточном, субклеточном и даже молекулярном уровнях является выдающейся, но не единственной отличительной чертой современной биологии. Для нее типичен углубленный интерес к процессам в сообществах организмов, которые определяют планетарную роль жизни. Таким образом, классификация пополнилась надорганизменными уровнями, такими, как видовой, биогеоценотический, биосферный. Разобранной выше классификации придерживается большинство конкретных медико-биологических и антропобиологических наук. Это неудивительно, так как она отражает уровни организации живой природы через исторически сложившиеся уровни ее изучения. В задачу курса биологии медицинского вуза входит преподать наиболее полную характеристику биологического «наследства» людей. Для решения этой задачи целесообразно воспользоваться классификацией, наиболее близко отражающей именно уровни организации жизни. В названной классификации выделяются молекулярно-генетический, клеточный, Организменный, или онтогенетический, популяционно-видовой, биогеоценотический уровни. Особенность данной классификации заключается в том, что отдельные уровни иерархической системы жизни определяются в ней на общей основе выделения для каждого уровня элементарной единицы и элементарного явления. Элементарная единица — это структура или объект, закономерные изменения которых, обозначаемые как элементарное явление, составляют специфический для соответствующего уровня вклад в процесс сохранения и развития жизни. Соответствие выделяемых уровней узловым моментам эволюционного процесса, вне которого не стоит ни одно живое существо, делает их всеобщими, распространяющимися на всю область жизни, включая человека. Элементарной единицей на молекулярно-генетическом уровне служит ген — фрагмент молекулы нуклеиновой кислоты, в котором записан определенный в качественном и количественном отношении объем биологической (генетической) информации. Элементарное явление заключается прежде всего в процессе конвариантной редупликации, или самовоспроизведении, с возможностью некоторых изменений в содержании закодированной в гене информации. Путем редупликации ДНК происходит копирование заключенной в генах биологической информации, что обеспечивает преемственность и сохранность (консерватизм) свойств организмов в ряду поколений. Редупликация, таким образом, является основой наследственности. В силу ограниченной стабильности молекул или ошибок синтеза в ДНК (время от времени, но неизбежно) случаются нарушения, которые изменяют информацию генов. В последующей редупликации ДНК эти изменения воспроизводятся в молекулах-копиях и наследуются организмами дочернего поколения. Указанные изменения возникают и тиражируются закономерно, что и делает редупликацию ДНК конвариантной, т.е. происходящей иногда с некоторыми изменениями. Такие изменения в генетике получили название генных (или истинных) мутаций. Конвариантность редупликации, таким образом, служит основой мутационной изменчивости. Биологическая информация, заключающаяся в молекулах ДНК, не участвует непосредственно в процессах жизнедеятельности. Она переходит в действующую форму, будучи перенесена в молекулы белков. Отмеченный перенос осуществляется благодаря механизму матричного синтеза, в котором исходная ДНК служит, как и в случае с редупликацией, матрицей (формой), но для образования не дочерней молекулы ДНК, а матричной РНК, контролирующей биосинтез белков. Отмеченное дает основание причислить матричный синтез информационных макромолекул также к элементарному явлению на молекулярно-генетическом уровне организации жизни. Воплощение биологической информации в конкретные процессы жизнедеятельности требует специальных структур, энергии и разнообразных химических веществ (субстратов). Описанные выше условия в живой природе обеспечивает клетка, служащая элементарной структурой клеточного уровня. Элементарное явление представлено реакциями клеточного метаболизма, составляющими основу потоков энергии, веществ и информации. Благодаря деятельности клетки поступающие извне вещества превращаются в субстраты и энергию, которые используются (в соответствии с имеющейся генетической информацией) в процессе биосинтеза белков и других соединений, необходимых организму. Таким образом, на клеточном уровне сопрягаются механизмы передачи биологической информации и превращения веществ и энергии. Элементарное явление на этом уровне служит энергетической и вещественной основой жизни на всех других уровнях ее организации. Элементарной единицей организме/того уровня является особь в ее развитии от момента зарождения до прекращения существования в качестве живой системы, что позволяет также назвать этот уровень онтогенетическим. Закономерные изменения организма в индивидуальном развитии составляют элементарное явление данного уровня. Эти изменения обеспечивают рост организма, дифференциацию его частей и одновременно интеграцию развития в единое целое, специализацию клеток, органов и тканей. В ходе онтогенеза в определенных условиях внешней среды происходит воплощение наследственной информации в биологические структуры и процессы, на основе генотипа формируется фенотип организмов данного вида. Элементарной единицей популяционно-видового уровня служит популяция — совокупность особей одного вида. Объединение особей в популяцию происходит благодаря общности генофонда, используемого в процессе полового размножения для создания генотипов особей следующего поколения. Популяция в силу возможности межпопуляционных скрещиваний представляет собой открытую генетическую систему. Действие на генофонд популяции элементарных эволюционных факторов, таких, как мутационный процесс, колебания численности особей, естественный отбор, приводит к эволюционно значимым изменениям генофонда, которые представляют элементарные явления на данном уровне. Организмы одного вида населяют территорию с известными абиотическими показателями (климат, химизм почв, гидрологические условия) и взаимодействуют с организмами других видов. В процессе совместного исторического развития на определенной территории организмов разных систематических групп образуются динамичные, устойчивые во времени сообщества — биогеоценозы, которые служат элементарной единицей биогеоценотического (экосистемного) уровня. Элементарное явление на рассматриваемом уровне представлено потоками энергии и круговоротами веществ. Ведущая роль в этих круговоротах и потоках принадлежит живым организмам. Биогеоценоз — это открытая в вещественном и энергетическом плане система. Биогеоценозы, различаясь по видовому составу и характеристикам абиотической своей части, объединены на планете в единый комплекс — область распространения жизни, или биосферу. Приведенные выше уровни отражают важнейшие биологические явления, без которых невозможны эволюция и, следовательно, само существование жизни. Хотя элементарные единицы и явления на выделяемых уровнях различны, все они тесно взаимосвязаны, решая свою специфическую задачу в рамках единого эволюционного процесса. С конвариантной редупликацией на молекулярно-генетическом уровне связаны элементарные основы этого процесса в виде явлений наследственности и истинной мутационной изменчивости. Особая роль клеточного уровня состоит в энергетическом, вещественном и информационном обеспечении происходящего на всех других уровнях. На онтогенетическом уровне биологическая информация, находящаяся в генах, преобразуется в комплекс признаков и свойств организма. Возникающий таким образом фенотип становится доступным действию естественного отбора. На популяционно-видовом уровне определяется эволюционная ценность изменений, относящихся к молекулярно-генетическому, клеточному и онтогенетическому уровням. Специфическая роль биогеоценотического уровня состоит в образовании сообществ организмов разных видов, приспособленных к совместному проживанию в определенной среде обитания. Важной отличительной чертой таких сообществ является их устойчивость во времени. Рассмотренные уровни отражают общую структуру эволюционного процесса, закономерным результатом которого является человек. Поэтому типичные для этих уровней элементарные структуры и явления распространяются и на людей, правда, с некоторыми особенностями в силу их социальной сущности.
Популярное:
|
Последнее изменение этой страницы: 2017-03-08; Просмотров: 837; Нарушение авторского права страницы