|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ОТДЕЛ БУРЫЕ ВОДОРОСЛИ — РНАЕОРНУТА
Для водорослей этого отдела характерны хроматофоры, окрашенные в бурый цвет благодаря тому, что помимо хлорофиллов «а» и «с» (хлорофилл «Ъ» отсутствует) и р-каротина имеется избыток бурых ксантофиллов,, особенно фукоксантина. Хлоропласты, или хроматофоры, помимо оболочки окружены сложной системой мембран, находящейся в прямой связи с оболочкой ядра — «хлоропластной эндоплазматической сетью». Матрикс хлоропласта пересекается параллельными трехтилакоидными ламеллами, окружёнными одной или несколькими периферическими (опоясывающими) ламеллами. Свободный от тилакоидов пиреноид выступает из хлоропласта в виде почки (рис. 152). Генофор кольцевидный, находится под перифери-
ческиыи лам. ми. Запасной полисахарид — ламинарии — откладывается вне хлоропласта в цитоплазме. Помимо ламинарина запасными продуктами служат шестиатомный спирт маннит и жиры. Монадные клетки (зооспоры и гаметы) имеют глазок и жгутики. Глазок — часть пластиды, связан со жгутиковым аппаратом. Жгутики, как правило, в числе двух (у диктиотовых — один), гетероконтные и гетероморфные. Мембрана переднего, обычно более длинного жгута на своей поверхности несет сложно устроенные масти- гонемы, которые состоят из трех частей: 1) базальной части (не трубчатой), которая прочно приклеивается к мембране жгута, 2) микротрубчатой части, образованной рядами глобулярных субъединиц, и 3) одной— трех терминальных нитей из одного ряда глобулярных субъединиц (рис. 153, А, В). Мастигонемы формируются внутри клетки в одетых мембраной пузырьках, по-видимому, отшнуровывающихся от перинук- леарного пространства (области между двумя мембранами ядерной оболочки). По мере образования мастигонемы выделяются на поверхность клетки вблизи места отхожде- ния переднего жгута, к мембране которого они и прикрепляются. О роли мастигонем пока ничего не известно. Задний (обычно более короткий) жгут гладкий, его основание, прилегающее к глазку, несет вздутие (рис. 153, Г). У гамет эктокар- пуса как передний жгут с трубчатыми мастигонемами, так и задний гладкий жгутик несут длинный, час- то спирально закрученный терминальный придаток — акронему, в которую из стержня жгута переходят две центральные микротрубочки (рис. 153, Л, Б). С помощью акро- немы переднего жгута мужской гаметы осуществляется начальный контакт ее с остановившейся и втянувшей свои жгутики женской гаметой. Сперматозоиды диктиотовых характеризуются лишь одним передним жгутом. За немногими исключениями бурые водоросли — морские, особенно богато представленные в холодных водах северного и южного полушария. По морфологической и анатомической дифференцировке таллома они стоят на более высоком уровне, чем ранее рассмотренные группы водорослей. Среди них неизвестны ни одноклеточные, ни колониальные формы, ни талломы в виде простой неразветвленной нити. Таллом самых простейших из ныцеживущих бурых водорослей гетеротрихальный, у громадного же большинства талломы крупные, компактные ложно- или истиннотканевого строения. Клетки имеют сильно ослизняющиеся стенки, содержат одно ядро, одну или много вакуолей, обычно постенные хроматофоры различной формы. Размножение вегетативное, бесполое и половое. Вегетативное размножение осуществляется участками таллома. У некоторых имеются специализированные веточки (выводковые почки), которые легко отламываются и вырастают в новые талломы. Бесполое размножение у большинства бурых водорослей происходит посредством зооспор, образующихся в одногнездных или однокамерных спорангиях, которые развиваются на диплоидных растениях (спорофитах) и в которых перед формированием зооспор яд{5о редукционно делится. У представителей порядка диктиотовых (01с1уо1а1е8) в одногнездных спорангиях (тетра- спорангиях) вместо зооспор, также после редукционного деления ядра, образуются четыре неподвижные апланоспоры — тетраспоры. Гаплоидные зооспоры и тетраспоры прорастают в гаплоидные половые растения — гаметофиты, на которых образуются • половые органы. У простейших бурых водорослей половой процесс изогамный, гаметы развиваются в многогнездных или многокамерных спорангиях, состоя- -> щих из большого числа (до нескольких сотен) мелких кубических клеток. Протопласт каждой клетки превращается в одну гамету. У ряда бурых водорослей наблюдается гетерогамия, причем гаметы также образуются в многокамерных спорангиях. Спорангии двух видов: одни состоят из большого числа мелких клеток, другие —из меньшего количества более крупных клеток. Соответственно в первых формируются мелкие микрогаметы, а во вторых — крупные макрогаметы. У наиболее высокоорганизованных бурых водорослей половой про-- цесс оогамный. В оогониях и антеридиях, как правило, образуется по одной гамете (яйцеклетке и сперматозоиду соответственно). Исключение составляют представители порядка фукусовые (Риса1е5), в антеридиях которых развиваются 64 сперматозоида, а оогонии у некоторых родов содержат более одной (две, четыре, восемь) яйцеклетки; например, у рода фукус [Рисиз) в оогонии образуется восемь яйцеклеток. Антеридии диктиотовых имеют вид многокамерных спорангиев, в каждой камере развивается по одному сперматозоиду, снабженному в отличие от монадных клеток всех остальных бурых водорослей только одним жгутиком. Яйцеклетка оплодотворяется всегда вне оогония. Зигота без периода покоя прорастает в диплоидное растение. У всех бурых водорослей, за исключением представителей порядка фукусовые, у которых отсутствует бесполое размножение и которые являются диплонтами, наблюдается смена генераций: у одних изоморфная, у других гетероморфная. Эти разные типы жизненного цикла положены в основу современного деления отдела бурых водорослей (РЬаеорЬу1а) на три класса, или когорты (группы порядков): изо- генератные (150^епега1ае), характеризующиеся изоморфной сменой генераций, гетерогенератные (Не1его^епега1ае), включающие бурые водоросли с гетероморфной сменой генераций, и ци& лоспоровые (Су- с1озрогеае) с одним порядком фукусовые, где смена генераций отсутствует. КЛАСС ИЗОГЕНЕРАТНЫЕ — 1$ООЕ^РАТАЕ Порядок эктокарповые — Ес1осагра1е$ Представителем порядка может служить род эктокарпус (Ес1осагри$, рис. 154), который объединяет, по-видимому, самые примитивные ныне- живущие бурые водоросли. Многочисленные виды его распространены во всех морях, особенно холодных, и растут на подводных предметах и других более крупных водорослях. Таллом (как спорофита, так и гаме- тофита) имеет вид желтовато-бурых кустиков, достигающих в длину нескольких сантиметров. Он состоит из стелющихся по субстрату нитей, от которых отходят вертикальные нити из одного ряда клеток, обильно ветвящиеся. Ветви вертикальных нитей часто оканчиваются многоклеточными бесцветными волосками. Стелющиеся по субстрату нити обнаруживают верхушечный рост. Рост вертикальных нитей у большинства видов диффузный, лишь у некоторых видов у основания волосков дифференцируется ясно выраженная интеркалярная (вставочная) меристема, еще реже встречается верхушечный рост.
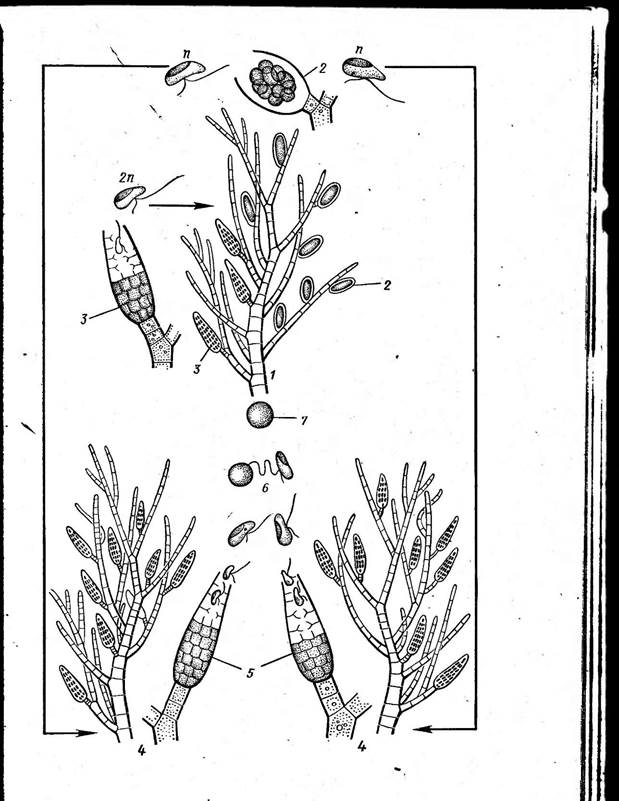
Бесполое размножение — зооспорами. Они развиваются на диплоидных растениях в одногнездных спорангиях, представляющих собой конечные клетки коротких боковых ветвей. Содержимое их распадается на ряд голых зооспор, которые выходят через разрыв стенки спорангия. Образованию зооспор предшествует редукционное деление ядра, и гаплоидные таким образом зооспоры вырастают в гаплоидные растения, размножающиеся половым путем. Гаметы формируются в многогнездных спорангиях. Хотя они внешне одинаковы и половой про-, цесс у эктокарпуса изогамный, по поведению различают женские и мужские гаметы. Первые быстро теряют подвижность, вторые могут оставаться подвижными до 24 ч. Остановившаяся женская гамета выделяет пахучее, сильно летучее вещество — углеводород эктокарпен (СиН16), привлекающий мужские гаметы, которые роем окружают женскую гамету и прочно прикрепляются к ее мембране с помощью акроне- мы переднего жгута. Затем одна из мужских гамет сокращает свой передний жгутик, приближаясь к женской гамете, с которой и сливается. Остальные мужские гаметы уплывают. Зигота без периода покоя прорастает в диплоидное бесполое растение. Таким образом, эктокарпус обнаруживает изоморфную смену генераций, от которой в зависимости от климатических условий могут быть отклонения. Так, в некоторых местообитаниях на спорофитах эктокарпуса помимо одногнездных спорангиев наблюдаются многогнездные — нейтральные спорангии, которые в отличие от многогнездных спорангиев, развивающихся на гаметофитах и являющихся гаметангиями, образуют в своих клетках зооспоры — нейтральные споры. Нейтральные споры диплоидны, так как перед их формированием редукционного деления ядра не происходит, они прорастают в диплоидные растения— спорофиты (рис. 155). Порядок сфацеляриевые — 5рНасе1апа1е$ Род сфацелярия (5рНасе1аг1а, рис. 156), как и эктокарпус, широко распространен во всех морях и имеет небольшие гетеротрихальные талломы. Однако рост как распростертых, так и вертикальных нитей в талломе сфацелярии строго верхушечный. Апикальная (верхушечная) с густым содержимым клетка делится поперечной перегородкой и последовательно отделяет вниз клетки-сегменты. Клетки-сегменты делятся, в свою очередь, один раз поперечно и каждая из двух дочерних клеток затем делится продольными перегородками, как радиальными, так и тангентальными. В результате этих делений более старые части тал- Рис. 155. Ес1осагри$. Цикл развития: на спорофите (/) могут возникать как одногнезд- ные (2), так и многогнездные (3) зооспорангии. В первых после мейоза образуются гаплоидные (п) зооспоры, во вторых — диплоидные (2п) зооспоры (нейтральные споры); гаплоидные зооспоры прорастают в гаметофиты (4), несущие многогнездные спорангии (гаметангии) (5). Гамегы (6) копулируют, зигота (7) прорастает в диплоидный спорофит (/). Диплоидные (нейтральные) зооспоры сразу прорастают с образованием диплоидного спорофита (1)
Рис. 156. 8р1юсе1апа. Кончик таллома с верхушечными клетками Рис. 157. 8рИасе1апа. Участок таллома с выводковыми почками
лома приобретают паренхиматозное строение. На поперечном разрезе через нижерасположенные части таллома в середине находится группа более крупных, бедных содержимым клеток, которые окружены корой из одного слоя мелких, богатых хроматофорами клеток. Р1екоторые более крупные клетки коры дают выступ, который.отчленяется перегородкой и начинает функционировать в качестве, верхушечной клетки боковой ветви. Так происходит ветвление таллома. Все сказанное относительно размножения и цикла развития экто- карпуса в полной мере приложимо к сфацелярии. Диплоидные бесполые растения некоторых видов сфацелярии на одном и том же индивиде могут образовывать как одногнездные, гак и многогнездные (нейтральные) спорангии. При морфологической изогамии сфацелярии ее гаметы различаются по поведению: мужские дольше остаются подвижными, и копуляция происходит только после того, как женские гаметы остановились. Для вегетативного размножения служат особые специализированные веточки — выводковые почки, которые отламываются и прорастают в новые талломы (рис. 157). Рис. 158. СНае1ор1еп$. Продольный разрез взрослого побега У рода хетоптерис (СНае1ор1ег1з, рис. 158), который, как и все сфацеляриевые, характеризуется етрого верхушечным ростом, в старых частях вертикальных нитей некоторые клетки первичной коры дают выросты, которые разрастаются в многоклеточные нити, оплетающие плотным и толстым слоем основную ось. Боковые ветви, отходящие от клеток первичной коры, проходят сквозь эту вторичную кору. Наиболее крупный представитель сфацеляриевых — род кладосте- фус (С1а^05(ерНиз), часто встречающийся в Черном море и достигающий 20 см в длину. На разветвленных основных осях, увенчанных верхушечными клетками, мутовками располагаются укороченные вет- ^Ви более простого строения. В длинных осях, как и у хетоптериса, образуется вторичная кора. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 381; Нарушение авторского права страницы
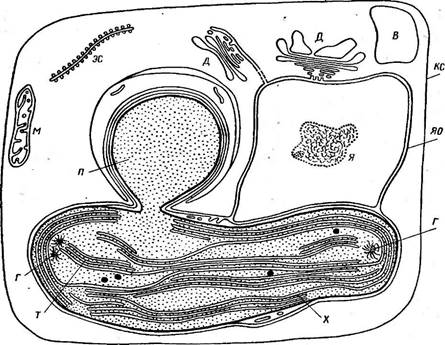 Рис. 152. Схема строения клетки бурой водоросли:
КС — клеточная стенка, « — митохондрии, ; > с —фрагмент канала эндоплазматической сети, д — диктио- семы, в — вай^оль, Я — ядро, яо — ядерная оболочка (в расширении между двумя ее мембранами — в пер ииу к л ео л яр ном пространстве — заключен хлоропласт), X — хлоройлаег с системой тилакоидов {7), собранных В пачки по три, п —пиреноид с обкладкой, г— кольцевидный генофор, перерезанный поперек
Рис. 152. Схема строения клетки бурой водоросли:
КС — клеточная стенка, « — митохондрии, ; > с —фрагмент канала эндоплазматической сети, д — диктио- семы, в — вай^оль, Я — ядро, яо — ядерная оболочка (в расширении между двумя ее мембранами — в пер ииу к л ео л яр ном пространстве — заключен хлоропласт), X — хлоройлаег с системой тилакоидов {7), собранных В пачки по три, п —пиреноид с обкладкой, г— кольцевидный генофор, перерезанный поперек
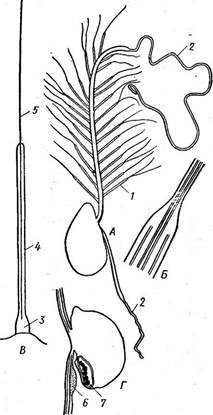 Рис. 153. Схема строения монадных клеток бурых водорослей. А — гамета Е& осагриз: передний жгутик с мастигонемами (/) и акронемой (2), задний жгутик гладкий, с акронемой (2); Б — переход от стержня жгута к ак- ронеме, в которую переходят две центральные микротрубочки аксонемы жгута; В —схема строения мастигонемы у АхорИуНит (3 — основные мастигонемы, 4 — трубчатая часть. 5 — терминальная нить); Г — сперма тозоид Рисиз со вздутием (6) у основания заднего жгута, прилегающего к стигме (7)
Рис. 153. Схема строения монадных клеток бурых водорослей. А — гамета Е& осагриз: передний жгутик с мастигонемами (/) и акронемой (2), задний жгутик гладкий, с акронемой (2); Б — переход от стержня жгута к ак- ронеме, в которую переходят две центральные микротрубочки аксонемы жгута; В —схема строения мастигонемы у АхорИуНит (3 — основные мастигонемы, 4 — трубчатая часть. 5 — терминальная нить); Г — сперма тозоид Рисиз со вздутием (6) у основания заднего жгута, прилегающего к стигме (7)
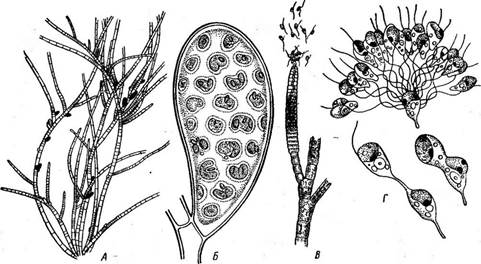 Рис. 154. ЕЫосагрш. А — общий вид; Б — зооспорангий (одногнездный спорангий) с зооспорами; В — гаметангий (миогогнездный спорангий) и гаметы; Г—оплодотворение
Рис. 154. ЕЫосагрш. А — общий вид; Б — зооспорангий (одногнездный спорангий) с зооспорами; В — гаметангий (миогогнездный спорангий) и гаметы; Г—оплодотворение