
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
КЛАСС ЦЕНТРИЧЕСКИЕ —СЕМКОРНУСЕАЕ
Виды класса особенно широко представлены в планктоне морей и океанов как одни из главных продуцентов органических веществ. Это одноклеточные или колониальные формы, характеризующиеся радиальной симметрией клеток, отсутствием активной подвижности (не имеют швов на панцире) и оогамным половым процессом. В классе содержится пять порядков, различающихся главным образом формой панциря и очертаниями створок. В планктоне и на дне пресных и морских водоемов обычны виды рода циклотелла (СусШеИа), иногда 'образующих колонии в виде непрочных цепочек. Клетки их похожи на круглую невысокую коробочку. Краевая зона створки несет радиальные штрихи или ребрышки, а центральная часть более выпукла и у большинства видов бесструктурна (рис. 146, А). Многочисленные хроматофоры в виде мелких пластинок располагаются по створковым сторонам. Сходный внешний вид имеют представители рода косцинодискус (Со$с1посИ$си8), встречающиеся в тех же местообитаниях. Но на их створковых сторонах радиальными или тангентальными рядами, реже беспорядочно располагаются точки, или ареолы, а по краю створки у некоторых видов имеются шипики (рис. 146, Б). Клетки в виде более высоких цилиндрических или бочонкообразных коробочек характерны для видов рода мелозира (Ме1о81га), обитающих преимущественно в планктоне и бентосе пресных и соленых водоемов. Клетки соединены створками в плотные нити (колонии) при помощи слизи, а у некоторых видов — и при помощи шипиков. Створки всегда круглые, покрыты нежными или грубыми точками (порами), часто образующими радиальные ряды. Многочисленные хроматофоры имеют вид дисков или лопастных образований (рис. 146, В). У видов рода хетоцерос (СИае1осего$) клетки иногда имеют вставочные ободки. Особенно же типичны для них длинные полые щетинки или шипы на каждом из полюсов их эллиптических створок. Этими шипами они обычно соединяются в длинные цепочки, при этом шипы соседних клеток перекрещиваются и иногда срастаются в основании (рис. 146, Г). Эти виды почти исключительно морские, планктонные. Как уже говорилось, вегетативные клетки представителей класса
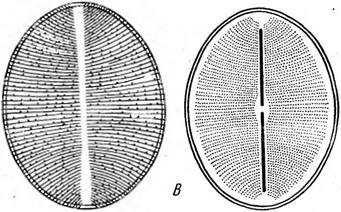
Созс/посИвсив, панцирь со створки;
В — МеШга: / — нить с пояска. 2 — иить с ауксоспорами; Г — СНае{осего5: / — клетка со створки, 2 — часть цепочки (клетки видны с пояска) центрические не обладают подвижностью, что, по-видимому, связано с жизнью в планктоне, но в то же время они эволюционировали в направлении усложнения строения клетки. Для них характерны различные выросты (хетоцерос), образование тонких малоокремневших промежуточных поясков, увеличивающих поверхность клетки и уменьшающих ее плотность, и другие признаки, носящие такой же приспособительный характер, например отложение в клетках масла, имеющего меньшую плотность, чем вода, мощные обвертки из слизи вокруг клеток, сравнительно слабое окремнение панциря и т. д. Последнее объясняется очень малым содержанием кремния в поверхностных слоях морей и океанов. КЛАСС ПЕРИСТЫЕ, ИЛИ ПЕННАТНЫЕ, — РЕШАТОРНУСЕАЕ Одноклеточные или колониальные водоросли. Клетки линейные или ланцетовидные, реже эллиптические или округлые, биполярные, с перистой структурой панциря. Среди них есть подвижные (со швом) и неподвижные. Половой процесс типа конъюгации. Класс содержит четыре порядка, различающихся в основном степенью развития шва (от бесшовных до обладающих каналовидным швом). Типичные представители порядка шовные (КарЫпа1е8) — виды рода пиннулярия (Ригпи1апа), встречающиеся на дне или в обрастаниях у берегов в различных, преимущественно пресных водоемах. Предпочитают воды, бедные известью. Со створки эти одноклеточные водоросли имеют вид вытянутого эллипса. Вдоль етворковой стороны посередине проходит шов в виде тонкой, слегка изогнутой линии. На обоих концах клетки и в середине находятся терминальные и центральный узелки. Края створки имеют четкий рисунок из параллельных ребрышек, не доходящих до линии шва, которые представляют собой перегородки узких поперечных камер на внутренней стороне створки. Вдоль обоих краев створки Видны в профиль (так как размещаются на поисковых сторонах) крупные пластинчатые хроматофоры в виде двух узких полосок желто-бурого цвета. В центре клетки находится цито- плазменный мостик, в котором лежит ядро. По обе стороны от мостика имеются вакуоли, капли масла и зерна волютина. С пояска клетка имеет вид продолговатого прямоугольника и вся окрашена в желто-бурый цвет. В этом положении в ней видны все указанные органеллы и включения, но пояски панциря не несут никаких скульптурных утолщений и на загибе створки видны только конечные участки перегородок между камерами. Узелки имеют вид сосочкообраз- ных утолщений панциря (рис. 147, Л—Г). Размножаются вегетативным делением. Виды рода навикула (Л^аь1си1а, рис. 148, А) встречаются в тех же местообитаниях, что и пиннулярия, но чаще, чем она, в солоноватых и морских водах. Многие из них очень похожи на виды пиннулярии и отличаются от них главным образом отсутствием камер в створках. Кроме того, у многих видов концы клеток сужены таким образом, что 13 Заказ 552
форма клетки напоминает лодочку (от лат. пау1си1а — лодочка). Эпифитные виды рода навикула часто удерживаются на поверхности водорослей или высших растений, образуя на них полые слизистые трубки, в которых они свободно передвигаются и таким образоч выносятся над субстратом в более благоприятные условия освещения Виды сходных родов гиросигма (Сугоз^та) из пресных и плевг^- сигма (Р1еигоз1§та) из морских водоемов имеют створки, согн\ в виде буквы 5, и отличаются друг от друга штриховкой панциря (рис. 148, Б, В). Из подвижных бентосных форм также широко распространены в различных местообитаниях виды рода ницшия (ЫНгзсЫаУ. Клетки этих диатомей имеют палочковидные очертания, концы их заострены, на каждой створке имеется киль с каналовидным швом, идущий вдоль одного из краев створки (на противоположных сторонах у эпитекн и гипотеки, т. е. кили расположены по диагонали (рис. 148, Г). Очень массивны и сложны панцири у видов сурирелла (8ипге1- 1а)—еще одного рода из бентоса пресных, солоноватых и морсшг. водоемов. Со створки очертания ее эллиптические, яйцевидные, у некоторых гитаровидные, а с пояска клиновидные (рис. 148, Д). Вдо- каждой створки по краям проходят два гребня, или киля, с каналов; .- ным швом. Таким- образом, клетка здесь имеет всего четыре шва. Рис. 148. А — ЫиЫси1а, вид со створки; Б—Оуготдта, вид со створки; В — Р1еиго- та, вид со створки; Г—№7г«г/иа: 1 — клетка со створки, 2 — поперечное сечение; Д — ЗипгеНа: 1 — клетка со створки, 2 — клетка с пояска На дне литоральной зоны главным образом пресных водоемов и в обрастаниях широко распространены виды рода цимбелла (Сут- Ье11а). Створки их обычно полулунной, формы, с прямым или вогнутым •брюшным и выпуклым спинным краями. Шов обычно эксцентрический, более или менее приближенный к брюшному краю. Хроматофор один, расположен с поясковой стороны. Иногда клетки прикрепляются к субстрату слизистой ножкой, образующейся, на одном из концов, или заключены в студенистые трубки, где свободно перемещаются (рис. 149). В подобных же местообитаниях встречаются виды реда гомфонема (СотрНопета), в поперечной плоскости имеющие несимметричную форму (один конец клетки уже другого). Со створки клетка напоминает наконечник молотка, а с пояска клинообразна. Шов проходит посередине. Два хроматофора расположены по поисковым сторонам. У этих диатомей также может вырабатываться слизистая ножка, при этом возникают ветвистые колонии (рис. 150, А, Б). В обрастаниях водорослей из рода кладофора, вошерия или высших растений обычно присутствуют виды рода кокконеис (Соссопе1з). Клетки их имеют очертания эллипса и прикрепляются к субстрату всей плоскостью нижней створки, снабженной швом, а на верхней створке шов отсутствует (рис. 150, В). Хрома- тофор в виде подковообразно согнутой пластинки располагается на верхней створке. Из неподвижных перистых диатомей чаще других встречаются виды родов синедра (8упейга), фрагилярия (БгадИапа), табеллярия (ТаЬеИапа) и астерионелла (А81егюпе11а). Виды синедры обитают в литорали или обрастаниях пресных, солоноватых и морских вод (типичные планктонные формы также имеются, но очень редки). Они живут одиночно либо в виде пучковидно-вееровидных или звездчатых колоний, прикрепленных к субстрату или свободноплавающих. Клетки их палочковидны, н& концах заострены или закруглены и со стороны створок несут нежную поперечную штриховку. С пояска панцирь имеет прямоугольные очертания. Шва у них нет. Хроматофоры (два) располагаются в плоскости створок (рис. 151, А). У видов фрагилярии клетки очень напоминают клетки видов синедры, но соединяются створками в длинные лентообразные колонии, обитающие преимущественно в литорали, реже в планктоне пресных и солоноватых вод (рис. 151, Б). Клетки видов рода табеллярия имеют форму табличек (с пояска), снабжены вставочными ободками и септами и соединяются в колонии в виде зигзагообразной цепочки (реже колонии звездчатые). При этом клетки обращены друг к другу створковой стороной и соединяются своими уголками (слизью). Они характерны для литорали и планктона пресных вод (рис. 151, В). Колонии видов рода астерионелла, обитающие в планктоне пресных водоемов и морей, похожи на изящную звездочку с лучами (клетками), расположенными в разных плоскостях. Каждая клетка имеет вид тонкой палочки со слегка расширенными концами, причем свободные концы несколько уже, чем концы, которыми клетки соединяются в центре звездочки. Хроматофоры в клетках, так же как у видов рода табеллярия, имеют вид мелких зерен (рис. 151, Г).
Как уже указывалось, диатомовые водоросли широко представ-
лены в самых различных местообитаниях, часто встречаясь в массовых количествах. Однако некоторые виды и роды приурочены к специфическим экологическим условиям, определяемым химическим составом воды, освещенностью, степенью загрязнения органическими веществами й т. п. Так, виды рода мелозира — типичные представители планктона вод, богатых органической пищей, в то время как виды родов цикло-
телла, синедра встречаются в водах» бедных органическими веществами. При очень высокой степени эутрофности, вызываемой сточными водами, в планктоне появляются определенные популяции рода ницшия (АШшсЫа рака), и других родов. В кислых водах, богатых гумусовыми веществами, обнаруживаются, например, особые виды родов фрустулина (Ргиз(иИпа) и табеллярия. Специфическая флора диатомовых живет и в ручьях с чистой и холодной водой и т. п. В нижних течениях потоков с большим количеством органических веществ, где дно, камни и погруженные части водных растений часто покрыты слоем слизи, в ней в больших количествах скапливаются бактерии, микроскопические животные и водоросли, главным образом диатомовые. Обитатели слизи таких полисапробных зон все вместе образуют огромную физиологически активную адсорбирующую поверхность, играющую решающую роль в процессах самоочищения воды. Большое значение в распространении диатомовых водорослей имеют азот и фосфор, добавление которых обычно стимулирует их развитие. Этим объясняется, в частности, особенно большое скопление морских планктонных диатомей против устьев больших рек, приносящих воду, обогащенную азотом. Во влажном климате тропиков многие диа- томеи живут на листьях деревьев вместе с синезелеными водорослями, ассимилирующими азот атмосферы. Пресноводные диатомовые обнаруживают повышенную потребность в железе и хорошо развиваются при содержании Ре203 до 2—3 мг в 1 л воды. Многие диатомеи можно использовать как индикаторы водоема, а некоторые — даже как индикаторы морских течений. Так, в течении Гольфстрим доминирует один из видов центрической диатомовой водоросли— планктониелла (Р1апк1оп1е11а зо1, рис. 151, Д). В природе особенно большое значение имеют планктонные диатомовые. Составляя основную массу растительного планктона, они являются началом пищевой цепи. Их поедают беспозвоночные животные,, также обитающие в планктоне, которых в свою очередь поглощает рыбная молодь, а ее пожирают более крупные рыбы и другие животные. Некоторые взрослые рыбы и молодь питаются непосредственно диатомеями (сельдь, хамса, сардина и др.). Следовательно, мощное развитие диатомовых водорослей в планктоне имеет большое хозяйственное значение, так как им определяется богатство водоема рыбой. В литературе отмечается, например, что улов сардины на побережье Индийского океана зависит от развития одного из видов фрагилярии (РпщИапа осеатса). Аналогичное значение могут иметь и бентосные диатомеи. Установлено, что некоторые мелкие животные специализировались на питании определенными видами таких диатомей. Например, инфузории родов хилодон, окситрихия и другие питаются главным образом видами навикула и ницшия, потребляя по 30—40 экземпляров ежедневно. Обилием диатомей определяется и развитие личинок хиро- номпд. Однако массовое развитие некоторых диатомовых водорослей может иметь и отрицательное значение. Некоторые из них в больших количествах попадают в жабры личинок рыб и вызывают их гибель. Описан случай массового отмирания моллюсков, культивируемых в морской бухте Японии, жабры которых были буквально забиты планктонной диатомовой водорослью талассиозира (ТНа1аз8юз1га ёеар1епз). Иногда эти водоросли затрудняют лов рыбы, запутываясь в рыболовных сетях, что имело место, например, на Волге около Астрахани при массовом развитии мелозиры. У побережья Швеции то же явление вызвало огромное количество другой диатомеи — дидимосфении (й1йу- тозркаета §ет'та1а).... Большое практическое значение имеет горная мука, кизельгур, или диатомит, на 50—80 % состоящая из панцирей ископаемых диатомей, которые известны из отложений юрского периода. В палеогене, неогене и антропогенном периоде панцири планктонных диатомей, отлагавшиеся после отмирания водорослей на дне мелководных морских и пресноводных бассейнов, образовали там мощные отложения. Вынесенные на поверхность земли последующими горообразовательными процессами, они и сформировали указанную горную породу. Это масса белого или светло-серого цвета, очень легкая, пористая и твердая. Применяется как полировочный или шлифовальный материал, для тепловой и звуковой изоляции, как фильтрующее вещество в пищевой, медицинской и химической промышленности, ранее использовалась как связущая масса или наполнитель при изготовлении динамита. Знание ископаемых диатомовых водорослей помогает определить происхождение и возраст различных осадочных пород (так называемый диатомовый анализ). |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 324; Нарушение авторского права страницы

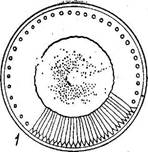
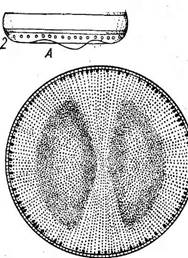
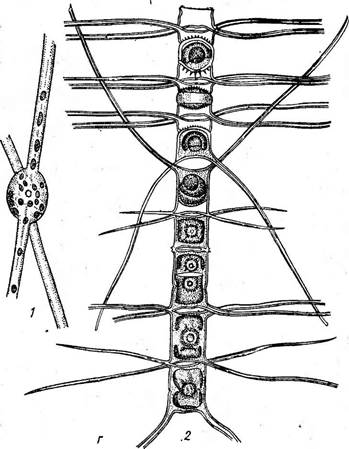
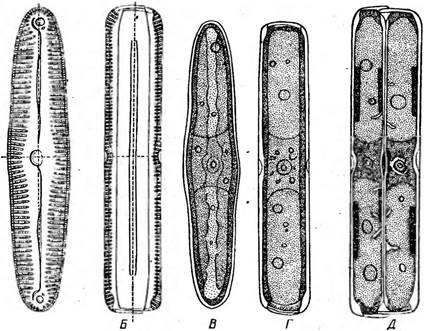 Рис. 147. Ртпи1апа. Л —панцирь со створки: Б — панцирь с пояска; В — клетка со створки; Г — клетка с пояска; Д—деление клетки (две дочерние клетки с пояска)
Рис. 147. Ртпи1апа. Л —панцирь со створки: Б — панцирь с пояска; В — клетка со створки; Г — клетка с пояска; Д—деление клетки (две дочерние клетки с пояска)
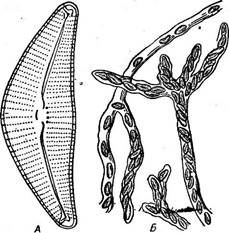 Рис. 149. СутЬеИа: А — панцирь со створки; Б—колония в студенистой трубке
Рис. 149. СутЬеИа: А — панцирь со створки; Б—колония в студенистой трубке
 1 I
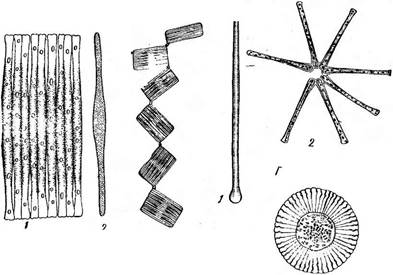
Рис. 150. А— Оотркопета:
/ — панцирь со створки. 2 — панцирь с пояска,
Б — колония Оотркопета-, В — Соссопеьв:
1 — верхняя створка, 2 — нижняя створка
1 I
Рис. 150. А— Оотркопета:
/ — панцирь со створки. 2 — панцирь с пояска,
Б — колония Оотркопета-, В — Соссопеьв:
1 — верхняя створка, 2 — нижняя створка
 А
Рис. 151. А — 8упеёга: ' / — панцирь со створки, 2 — панцирь с пояскаТ Б — РгадИагёа:
1 — колония, 2 — панцирь со створки;
В — ТаЬеНапа, общий вид колонии; Г — А$(епопе11а:
1 — панцирь со створки, 2 — общий вид колонии;
Д — Р1апк(рпСе11а зо1
А
Рис. 151. А — 8упеёга: ' / — панцирь со створки, 2 — панцирь с пояскаТ Б — РгадИагёа:
1 — колония, 2 — панцирь со створки;
В — ТаЬеНапа, общий вид колонии; Г — А$(епопе11а:
1 — панцирь со створки, 2 — общий вид колонии;
Д — Р1апк(рпСе11а зо1