|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ОТДЕЛ ДИАТОМОВЫЕ ВОДОРОСЛИ — С1АТОМЕАЕ, ВАС1ЬЬАРЮРНУТА
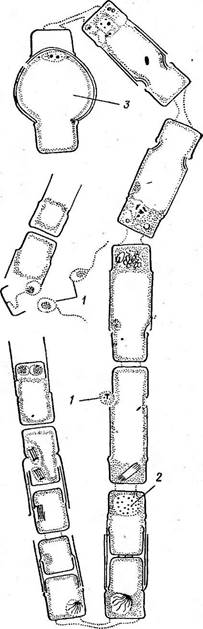
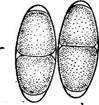
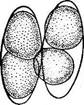
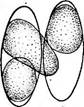
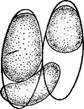
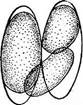
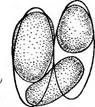
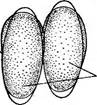
« Диатомовые водоросли — обширный отдел низших растений, чрезвычайно богатый видами (более 10 000). Это одноклеточные, и колониальные организмы микроскопически малых размеров, широко распространенные в бентосе и особенно в планктоне как морей и океанов, так и пресных вод с разным химическим составом и температурой. Некоторые виды обитают в верхних слоях почвы, на влажных скалах, в горячих источниках, на снегу и во льдах полярных областей. Ф. Нансен во время плавания на «Фраме» наблюдал появление на льду бурых пятен (скопления диатомей), под которыми лед быстро таял. Несмотря на большое разнообразие в строении отдельных форм и колоний им присущи общие специфические особенности в строении клетки и особенно клеточной оболочки. К наружному уплотненному слою протопласта клетки (плазмалемме) плотно примыкает панцирь из кремнезема (кремниевого гидрогеля, подобного опалу). Панцирь состоит из двух половинок, надеваю'щихся друг на друга, как крышка на коробку. Каждая половинка в свою очередь состоит из так называемой створки («донышка») и спаянного с ней поискового кольца. Большая створка— эпитека (крышечка) охватывает своим поисковым кольцом поисковое кольцо меньшей створки — гипотеки (коробочки). Очертания клетки зависят от ее положения, т. е. клетка может быть видна со створки, когда она лежит плашмя, или с пояска («в профиль»), когда видно, как пояски находят друг на-друга. У некоторых диатомей в течение жизни образуются дополнительные поясковые кольца или вставочные ободки между створкой и пояском. На них часто развиваются септы, или неполные перегородки, разделяющие' клетку на сообщающиеся между собой камеры. В зависимости от конфигурации клетки и структуры створок одни диатомовые имеют со стороны створки радиальную симметрию, другие— двустороннюю. Через первые можно провести ие менее трех плоскостей симметрии, а через вторые — не более двух. Первые выделяют в класс центрические (Сеп1горЬусеае), а вторые — в класс перистые, или пеннатные (Реппа1орЬусеае). Оболочка клетки не гомогенна. Во-первых, электронно-микроскопи- ческие и цитохимические исследования показали, что снаружи панциря, как и внутри него, располагается тонкий слой органического вещества. Во-вторых, на створках имеются характерные структуры. Они настолько постоянны и правильны, что имеют большое значение в систематике диатомей. Под микроскопом они видны как система точек, штрихов, ребер, ячеек и т. п. На самом же деле это или сквозные поры, или камеры, открывающиеся внутрь либо наружу, или чередующиеся более толстые и тонкие участки панциря. Их строение имеет разную степень сложности, а дно пронизано многочисленными отверстиями. Отверстия в стенке панциря, сообщающие протопласт клетки с окружающей средой, занимают от 10 до 75 % площади створки. Для многих планктонных форм характерны выросты на панцире. Клетки некоторых колониальных диатомей соединяются друг с другом при помощи этих выростов (см. рис. 146, 7" ). Некоторые бентоеные диатомеи из класса перистые способны к активному движению. Их содержимое сообщается с внешней средой продольной щелью или швом, проходящим или непосредственно по створ- ковой стороне, или по особым выростам панциря — килям, .расположенным вблизи длинной оси створки или по краю ееС движением связаны и так называемые узелки, или внутренние утолщения стенок створки, видимые под световым микроскопом как три небольшие круглые выпуклости: одна в центре и по одной на концах створковой стороны панциря (см. рис. 147, А, Б). ■ Движение диатомей — сложный процесс, ~ представляющий собой следствие перемещений цитоплазмы в шве и вертикальных каналах, проходящих в узелках. В движении принимают участие две системы орга- нелл: до 30 мелких, преломляющих свет частиц, расположенных вблизи шва в каждом конце клетки, и длинные фиброзные тяжи, или ленты, проходящие под швом. Светопреломляющие частицы выделяют локомоторное вещество через конечные поры и по всей длине шва; благодаря сокращению фиброзных тяжей локомоторное вещество выделяется, оставляя на субстрате слизистый след. Цитоплазма располагается в клетке тонким постенным слоем, а у некоторых и в середине клетки, где помещается ядро. Остальная, большая часть клетки заполнена вакуолью. Хроматофоры у большинства постенные, бывают в разном количестве (один, два, много). Это или крупные пластинки, иногда с пиреноидом, или зернистые образования часто неправильной формы. Они имеют желтую или желто-бурую окраску и содержат хлорофиллы «а» и «с», р- и 2-каротин, пять ксантофиллов (например, диадино-ксантин, в большом количестве фукоксантйн и др.). Мертвые клетки диатомей зеленеют, так как дополнительные к хлорофиллу пигменты разрушаются и выходят из клетки.. В ультрамикроскопическом строении хроматофора диатомей много общего с золотистыми водорослями; в частности, ламеллы состоят из трехтилакоидных дисков, имеется один опоясывающий тнлакоид. Продукты ассимиляции — масла, отлагающиеся в клетке в виде капель, волютин (или метахроматин) и хризоламинарин. У большинства диатомей ядро помещается в цитоплазменном мостике в центре клетки, у немногих располагается вблизи одной из. створок, имеет одно или несколько ядрышей и мелкие хроматиновыё зерна. У некоторых диатомей по обе стороны от ядра находится пара диктио- сом (аппарат Гольджи), обнаруживаемых под электронным микроскопом. Размножаются диатомеи, во-первых, вегетативным делением клеток, особенно интенсивным весной или в начале лета. Во-вторых, у них есть половой процесс. При вегетативных делениях масса протопласта увеличивается, вследствие чего обе половинки панциря отодвигаются друг от друга. Ядро митотически делится, затем протопласт разделяется пополам в плоскости, параллельной створкам. Каждый новый протопласт наследует половину панциря, а вторая образуется заново, причем у обеих дочерних клеток она будет меньшей — гипотекой 1 Шов может быть щелевидным, в виде узкой короткой или длинной щели, прорезывающей стенку створки, илн более сложным — каналовидным, расположенным в вы- •росте стенки створкн (в киле). С внешней средой он сообщается щелью, а в полость клетки открывается рядом отверстий с окремневшнми перегородками. рис. 147, Д). После этого две дочерние клетки у " одиночно живущих форм расходятся, а у колониальных остаются соединенными своими створками. Так как окремневшие панцири клеток не способны растягиваться, то при каждом делении одна из дочерних клеток равна материнской (у которой эпитека переходит от материнской клетки), а другая становится меньше (у которой эпитекой служит гипотека материнской клетки). В результате последующих делений размеры клеток в популяции прогрессивно уменьшаются. Математически ход деления диатомей можно выразить следующим образом: после п делений из одной клетки образуется 2П клеток и п+1 вариантов размеров. В каждый данный момент количество клеток каждого размера может быть выражено коэффициентом суммы биномов (х+у)п. Например, после четвертого деления общее количество клеток будет 24, т. е. 16. Вариантов размеров при этом будет 4+1=5, а количество клеток каждого размера — 1, 4, 6, 4, 1. При этом должно быть очень быстрое мельчание клеток в популяции, что иллюстрируется следующей таблицей (табл. 4).
В действительности деление диатомовых водорослей как в природе, так и в условиях культивирования протекает не в точном соответствии " с этим законом. Так, у некоторых форм, имеющих толстые панцири, например у видов рода мелозира (Ме1о8иа), можно наблюдать повторное деление более крупных дочерних клеток, в то время как меньшие не делятся. У некоторых диатомовых пояски панциря более эластичны, так что разница в величине эпитёки и гипогеки становится едва различимой. Электронно-микроскопическое изучение панцирей некоторых диатомей показало, что некоторые части панциря не срастаются, а сочленяются между собой подвижным механизмом, напоминающим сустав, что делает возможным раздвигание частей панциря и выравнивание разницы в размерах эпитеки и гипотеки. Обычно уменьшению размеров клеток диатомей в ходе последовательных делений противопоставляется увеличение их в результате полового процесса. Однако этот процесс происходит и у неизмельчавших клеток, поэтому можно предположить, что половой процесс зависит не только от уменьшения размеров клеток, но и от каких-то других факторов.
Б Рис. 145. А — схема полового процесса и образования ауксоспор (/) пеннатных диатомей на примере СотрНопета рамиШт; Б — Ме1оЫга: 1 — развитие сперматозоидов, 2 — развитие яйца, 3 — образование ауксоспоры У пеннатных диатомей {например, у гомфонемы) половой процесс напоминает конъюгацию десмидиевых водорослей (рис. 145, А). Две особи сближаются и выделяют слизь. В каждой клетке ядро редукци- онно делится на четыре, из которых у одних видов три, а у других два дегенерирует. В первом случае весь протопласт каждой клетки становится гаметой, во втором — образует две гаметы. Копуляция происходит таким образом, что гамета.одной из клеток, двигаясь амебообраз- но, переходит к гамете другой клетки, остающейся на месте. В случае ■ формирования каждой клеткой двух гамет одна из них переходит в копулирующую клетку, а другая остается на месте и сливается с гаметой, переползающей из другой клетки. У некоторых диатомей конъю- гирующие клетки, лежащие далеко друг от друга, могут образовать слизистые каналы для прохождения гамет. Подвижные гаметы можно рассматривать как мужские, а остающиеся на месте — как женские. Таким образом, при половом процессе этого типа можно говорить о физиологической анизогамии. В результате полового процесса, происходящего очень быстро (в течение нескольких минут), развиваются одна или две зиготы, которые увеличиваются в размерах и превращаются в так называемые ауксоспоры (от греч. «ауксо» — увеличиваю, расту). При этом тонкая оболочка зиготы расплывается или разрывается и ее остатки заметны на концах ауксоспоры. Зрелые ауксоспоры одеваются оболочкой, постепенно приобретающей характерную для данного вида структуру, и превращаются в вегетативные клетки. У некоторых диатомей ауксоспоры могут возникать в пределах одной клетки. При этом материнская клетка или образует две гаметы, копулирующие друг с другом, илц в ее неподелившемся протопласте сливаются два ядра (автогамия)/У некоторых имеет место диплоидный партеногенез, т. е. без всякого слияния клеток или ядер из вегетативной клетки формируется азигота, превращающаяся в ауксоспору. У центрических диатомей (например, у Ме1оз1га) обнаружен оогам- «ый половой процесс. В одних клетках образуются четыре сперматозоида с одним или двумя жгутиками, в других также происходит редукционное деление ядра, в результате которого в клетке остается только одно жизнеспособное ядро. Эта клетка соответствует оогонию с одной яйцеклеткой. Свободноплавающие сперматозоиды проникают в оогоний и оплодотворяют ее. Зигота одевается пектиновой оболочкой и превращается в ауксоспору (рис. 145, Б). Ийтересно, что оогамия доказана и у одного вида рода рабдонема (КкаЬкопета айпаИсит) из класса пеннатных диатомей. Таким образом, эта бесшовная диато- мея может рассматриваться как связующее звено между классами пеннатные и центрические. Как видно из обзора полового процесса диатомей, вегетативные особи представляют собой диплоидные организмы, а гаметы — гаплоиды. Это свидетельствует о том, что они проделали своеобразный и сложный путь эволюционного развития, не превратившись, однако, чв многоклеточные формы. Существование сперматозоидов у центрических диатомей и образование пульсирующих вакуолей при вегетативном делении у некоторых родов.(АНкеуа, НЫго5о1ета, Ш(гяск1а и др.) указывают на возможное происхождение диатомей от жгутиковых. В свою очередь жгутиковая стадия и палеонтологические данные свидетельствуют о том, что представители класса центрические должны рассматриваться как более примитивные и более древние, чем представители класса пеннатные. У некоторых морских и реже пресноводных представителей центрических диатомей известны цисты. Они очень похожи на цисты золотистых водорослей, что служит одним из доказательств филогенетической близости обоих отделов. Известно, что диатомеи могут долго существовать без воды. По-видимому, протопласт их способен к перенесению неблагоприятных условий в состоянии анабиоза. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 311; Нарушение авторского права страницы