|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Порядок ламинариевые — Ьаттапа1е5
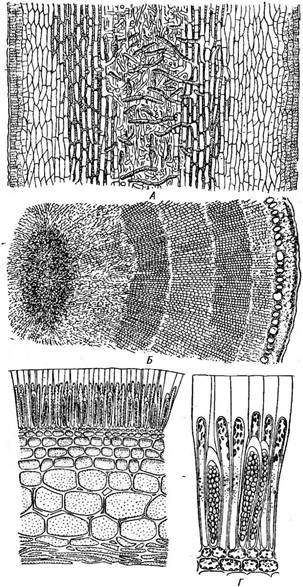
Гаметофиты всех представителей порядка мало различаются между собой и представлены микроскопическими, часто редуцированными до нескольких клеток нитчатыми растеньицами-заростками, которые несут половые органы (рис. 164). На мужских гаметофитах образуются антеридии в виде мелких клеток, развивающих по одному сперматозоиду, на женских — оогонии, в которых формируется по одной яйцеклетке. Последняя выходит из оогония и оплодотворяется вне его, после чего сразу прорастает в спорофит (рис. 164)) Спорофиты разных родов рез'ко различаются и представляют собой самые крупные низшие растения, достигающие в длину 60—100 м, со значительным морфологическим расчленением и сложным анатомическим строением. В большинстве случаев спорофиты ламинариевых расчленяются на листовидную пластинку, «ствол» и ризоиды, с помощью которых все растение прикрепляется к подводным камням и скалам. В месте перехода листовой пластинки в стволик находится интеркалярная меристема, за счет деятельности которой нарастает как листовая пластинка, так и ствол. У многолетних представителей зимует ствол с ризоидами, а листовая пластинка ежегодно отмирает и заменяется новой. В истинной ткани, образующей спорофиты ламинариевых, можно различать мелкоклеточную наружную кору, под которой располагается крупноклеточная внутренняя кора, граничащая изнутри с сердцевиной. Сердцевина образована главным образом рыхлым сплетением нитей (рис. 165, А, Б). В коре многих ламинариевых имеются особые ситовидные трубки с поперечными перегородками, пронизанными многочисленными отверстиями. Они, несомненно, выполняют проводящую функцию. Представителем порядка может служить род ламинария (Ьшт- пагьа), виды которого широко распространены в северных морях. Таллом спорофита, достигающий нескольких метров в длину, расчле- сэсэоез
нен на листовую пластинку, ствол и ризоиды (рис. 164, 7; 166, А). I Ствол и ризоиды многолетние, листовая пластин- на меняется ежегодно. При размножении на поверхности листовой пластинки из коровых клеток группами (сорусами) образуются одногнездные спорангии, в которых развиваются зооспоры (рис. 164, 2, 3, 165, В, Г). Зооспоры прорастают в микроскопически мелкие заростки (гаметофиты). Из, более крупных представителей порядка можно назвать роды мак- роцистис (МасгосузИз) и нереоцистис (Мегеосуз- /15). Талломы макроцис- тиса, распространенного в южном полушарии, достигают длины 60 м, имеют длинный тонкий (до 1 см в диаметре) ствол, который в нижней части твлый, а в верхней односторонне образует листья, снабженные каждый в основании воздушным пузырем (рис. 166, Б). Рис. 164. Ьаттапа. Цикл развития: на спорофите (/) развиваются сорусы зооспоран- гиев (2), в которых после редукционного деления образуются зооспоры (3). Они вырастают в микроскопические гаметофиты (4) — мужские с антеридиями (5) и женские с обгон ия ми (6). Оплодотворенная сперматозоидом (7) яйцеклетка (в) немедленно прорастает с образованием спорофита (/) Рис. 166. Ьагтпапакв. Спорофиты: А — Ьапипапа, Б — МасгосувИв, В — Иегеосу& И, Г — А1апа У нереоцистиса (рис. 166, В) длинный (до 25 м) сравнительно тонкий первичный ствол на верхушке вздувается в крупный воздушный пузырь, от которого отходят листовые пластинки (до 5 м длины), каждая с коротким вторичным стебельком. У макроцистиса, нереоцистиса, как и у ламинарии, зооспорангии располагаются в сорусах на листовых пластинках. В отличие от цих у рода алария (А1апа) сорусы зооспорангиев развиваются на специальных листочках — спорофиллах. Таллом аларии расчленен- на ствол с ризоидами и листовую пластинку с хорошо заметной срединной жилкой. От ствола отходят многочисленные спорофиллы (рис. 166, Г). КЛАСС ЦИКЛОСПОРОВЫЕ —СУСЬ08Р0КЕАЕ Этот класс представлен одним порядком фукусовые (Риса1е$). Порядок фукусовые — Риса1е5 Порядок характеризуется верхушечным ростом таллома, отсутствием бесполого размножения и оогамйым половым процессом, причем половые органы располагаются в углублениях таллома — концептаку- Рис. 167. Внешний вид фукусовых. А— 8аг/> аввит, Б — Рисив, В — СувШека лах, или скафидиях[18]. Не достигая, как правило, таких больших размеров, как представители ламинариевых, фукусовые вполне могут соперничать с ними по степени дифференцировки своего таллома. Наи- олее сложное морфологическое расчленение таллома наблюдается у ода саргассум (5агразвит), широко распространенного в южном Полушарии, особенно в теплых морях (рис. 167, Л). Стволик, прикрепленный к субстрату подошвой, моноподиально ветвится и несет уплощенные листовидные образования, шаровидные воздушные пузыри на- специальных стебельках и особые разветвленные плодущие веточки — Б Рис. 168. Рисив. А — разрез женского скафидия; Б — разрез мужского скафидия рецептакулы, на которых располагаются половые органы. Последние погружены в грушевидные углубления — концептакулы (скафидии), сообщающиеся с наружной средой через узкое отверстие. В оогонии образуется одна яйцеклетка, а в антеридии — 64 сперматозоида. Формированию половых клеток предшествует редукционное деление ядра. При созревании окруженные слизью половые клетки выходят из отверстия скафидия, и оплодотворение яйцеклетки происходит в окружающей воде. Зигота немедленно прорастает в диплоидный таллом. Как и у всех фукусовых, бесполое размножение у саргассума отсутствует. Имеется вегетативное размножение, приводящее в некоторых участках океана к образованию громадных скоплений талломов саргассума (примером может служить Саргассово море). В северных морях широко распространен род фукус (Рисиз) —■ основной обитатель береговой зоны (рис. 167, Б). Плоский ремневид- ный, дихотомически разветвленный таллом темно-бурого цвета достигает 1 м длины. Вдоль лопастей таллома с гладкими или зазубренными краями проходит срединная жилка, в нижней части переходящая в черешок, который прикрепляется к субстрату расширенным основанием. У некоторых видов фукуса по бокам от срединной жилки расположены вздутия, наполненные воздухом, — воздушные пузыри, у других видов они могут отсутствовать. Таллом нарастает благодаря деятельности верхушечных клеток, расположенных на концах плоских разветвлений. При размножении концы таллома вздуваются, принимают более светлую желто-оранжевую окраску и превращаются в рецептакулы, на которых образуются концептакулы — скафидии (рис. 168, А, Б). В. оогонии фукуса формируются восемь яйцеклеток, , в антери- диях — 64 сперматозоида. Яйцеклетки фукуса (Р. 8егга(и8), как и женские гаметы эктокарпуса, кутлерии, выделяют летучее вещество, привлекающее сперматозоиды. Это- вещество (его формула СвН12) названо фукосерратеном. В Черном море распространен род цистозейра (Су$1о5е1га) (см. рис. 167, В) с крупным (до нескольких метров) моноподиально разветвленным талломом, прикрепленным к камням подошвой. В нем обычно имеются воздушные пузыри. Веретеновидные рецептакулы несут скафидии. * * * Бурые водоросли — естественная группа, по своей морфологии хо^ рошо отличающаяся от остальных отделов водорослей. Однако, как уже говорилось, по ряду признаков (наличию хлорофиллов «а» и «с», отсутствию хлорофилла «Ь», расположению тилакоидов в хлоропластах — трехтилакоидные ламеллы, по сходным продуктам запаса — ламинарии, хризоламинарин, по строению монадных клеток с гетероконтными и гетероморфными жгутиками) бурые водоросли обнаруживают сходство с золотистыми, желтозелеными/диатомовыми и пирофитовыми водорослями. На этом основании некоторые авторы низводят бурые и другие перечисленные группы водорослей до ранга классов в пределах одного большого отдела СЬгошорЬу1а. Существование монадных клеток в жизненном цикле бурых водорослей позволяет выводить их, так же как остальные перечисленные группы, из каких-то первичных фото- синтезирующих жгутиконосцев с преобладанием бурых пигментов. Эти жгутиковые эволюционировали несколькими путями, один из которых и привел к возникновению бурых водорослей. Однако у бурых водорослей связь со жгутиковыми более отдаленная, чем у сходных с ними по ряду признаков золотистых, разножгутиковых, диатомовых, пиро- фитовых, - а также у зеленых водорослей, так, как прямых переходных форм здесь нет. Среди бурых водорослей не прослеживается последовательное развитие типов организации таллома от монадного к нитчатому и пластинчатому, как это легко наблюдается у зеленых, желтозе- леных, золотистых и пирофитовых водорослей. Как уже отмечалось, среди бурых водорослей представлены только высшие ступени морфологической дифференцировки таллома — гетеротрихальная и пластинчатая. Возможно, более простые формы (монадные, коккоидные, простые нитчатые) полностью утрачены на протяжении длительной эволюции, которую проделали бурые водоросли — древняя группа, известная уже из отложений силура и девона. Относительно родственных связей порядков в пределах отдела бурых водорослей и расположения их в системе единого взгляда нет. Согласно одной из схем филогенеза бурых водорослей, построенной с учетом сходства строения таллома на самых ранних стадиях развития, бурые водоросли — группа дифилетическая: одна линия эволюции объединяет (из упомянутых в изложении) сфацеляриевые, диктиотовые и фукусовые, другая — эктокарповые, кутлериевые, ламинариевые. Обе группы порядков различаются и по способу роста талломов: первая характеризуется верхушечным ростом, вторая — интеркалярным. Килин (Н. КуНп), система которого наиболее распространена в настоящее время, от общего предка бурых водорослей выводит три эволюционные линии, которые представлены им как классы изогенератные, гетеро- генератные и циклОспоровые; в основу их положены различия в жизненных циклах и морфологическом строении водорослей. Крупные бурые водоросли находят большое применение в хозяйстве человека. Из них добывают альгин — клеящее вещество, использующееся в текстильной, пищевой и ряде других отраслей промышленности. В приморских странах выбросы морских водорослей, богатые калием и азотом, употребляют в качестве удобрения, а также используют на корм скоту. Некоторые, например ламинарии («морская капуста»), съедобны. ОТДЕЛ ПИРОФИТОВЫЕ ВОДОРОСЛИ — РУКРОРНУТА Относящиеся к этому отделу в большинстве своем монадные водоросли, реже амебоидной, коккоидной, пальмеллоидной и нитчатой структуры, очень своеобразны по строению, так как совмещают признаки растений и животных. В их клетках, у многих одетых твердой оболочкой, содержатся хроматофоры, окрашенные преимущественно в бурый, темно-коричневый, красноватый и оливковый цвета, и крахмал в качестве продукта ассимиляции; вместе с тем у некоторых из них имеются щупальца, псевдоподии, стрекательные капсулы и характерный для животных тип питания. Общие для всех признаки — дорзовентральное строение монадных клеток, наличие бороздок на их поверхности и глотки. В составе отдела два класса: криптофитовые (Сгур1орЬусеае) и динофитовые (ОторНусеае) |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 311; Нарушение авторского права страницы