
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ОТДЕЛ ЛИШАЙНИКИ —ЫСНЕМОРНУТА
Лишайники — своеобразная группа организмов, в теле которых сочетаются два компонента: автотрофный фикобионт (водоросль или гонидии, как их называли ранее) и гетеротрофный микобионт (гриб), образующие единый симбиотический организм, отличающийся морфолого-анатомическп, физиолого-биохимическн, экологически, а следовательно, и качественно от свободноживущих грибов и водорослей. Вегетативное тело лишайников — слоевище (таллом), как и у других низших растбний, не дифференцировано на листья, стебель и корень. Окраска слоевища лишайников обусловлена различными пигментами и может быть серой, сизой, зеленоватой, буро-коричневой, желтой,, оранжевой, почти черной и др. Лишайники часто смешивают со мхами, но они отличаются от них отсутствием типичной зеленой окраски и диф- ференцировкой слоевища на органы. Двойственная природа лишайников открыта в 60-х годах XIX в. немецким ботаником С. Швенденером. Доказательством такой природы служат: 1) морфолого-анатомическое строение лишайников, показывающее, что фикобионты лишайников являются водорослями, а бесцветные нити между ними — грибными " гифами; 2) отсутствие генетических связей между фикобионтами и микобионтами; 3) возможность изолирования водоросли и гриба в чистую культуру; 4) получение на синтетических средах слоевищ лишайников из спор, взятых из спороношений гриба лишайников, и чистых культур соответствующих водорослей. \ Взаимоотношения гриба и водоросли в лишайнике В настоящее время сложились три мнения о характере взаимоотношений симбионтов лишайника: 1) паразитизм гриба на водоросли, 2) ило- тизм, 3) мутуалнстическое сожительство. Первое мнение основывается на наблюдениях в световой и электронный микроскопы, показавшие, что гриб образует апрессории,. давящие на клеточную стенку водоросли, и гаустории, проникающие- внутрь клеток фикобионта и приводящие их к гибели. Образование гаусторий не является облигатным и заметно лишь у некоторых клеток. В слоевище лишайника постоянно отмирают клетки водоросли как под. влиянием апрессорий и гаусторий гриба, так и в процессе онтогенеза фикобионта без взаимодействия с микобионтом. Отмершие клетки заметны и в сердцевине слоевища, и в коровом слое. Иногда видна не- кральная (отмершая) зона, что внешне проявляется в изменении цвета коры лишайника. При таких взаимоотношениях гриб может питаться паразитически за счет живых клеток фикобионта и Сапротрофно, употребляя отмершие клетки и продукты их обмена, находящиеся в слизистых выделениях клеток. На возможность двойного способа питания грибов в лишайниках в 30-х годах настоящего столетия обратили внимание русские ученые А. А. Еленкин и А. Н. Данилов, предложившие называть взаимоотношения гриба и водоросли в лишайнике эндопара- зитосапрофитизмом. Позднее было установлено, что и фикобионт может иногда переходить к миксотрофному питанию, потребляя разлагающиеся грибные гифы. В то же время микобионт, тесно оплетающий клетки фикобионта абсорбционными органами (апрессориями и гаусториями), истощает их, усваивая продукты метаболизма. Фикобионты лишайника лишены таких обычных (в зависимости от систематической принадлежности) ассимилятов, как крахмал, липиды, зерна цианофицина и др. Имеются прямые наблюдения о движении углеводов между компонентами лишайника. Установлено, что в лишайниках с синезелеными фикобионтами гриб потребляет глюкозу, у видов с зелеными фикобионтами — многоатомные спирты (рибит или эритрит, или сорбит). Поглощение того или иного спирта зависит от рода фикобионта, являющегося компонентом лишайника. Исследования метаболизма азота показали, что способностью по- тлощать связанный и свободный азот обладают преимущественно водоросли. Атмосферный азот фиксируется синезелеными фикобионтами -слоевища лишайника. Зеленые фикобионты поглощают связанный азот и наиболее доступная его форма — соли аммония. В 30—40-х годах XX в. советский ученый П. А. Генкель и независимо от него итальянская исследовательница Ченджа Самбо обнаружили в слоевище некоторых лишайников азотобактер — бактерию, способную усваивать атмосферный азот, и приняли его за третий симбионт лишайников. Аминокислоты, образующиеся в процессе метаболизма фиксированного азота, служат дополнительным источником азотного питания тех лишайников, в которых имеется азотобактер. Однако при-~ сутствие азотобактера в лишайниках не облигатно, необязательно, поэтому его нельзя считать третьим компонентом лишайников. , К теории паразитизма близка теория илотизма. Согласно этой теории гриб в слоевище лишайника играет роль регулирующего «хозяина», «эксплуатирующего» водоросль, но создающего условия, при которых фикобионт продолжает жить и размножаться. Действительно, гетеротрофный компонент (микобионт) использует органические вещества, создаваемые автотрофным компонентом (фикобионтом) умеренно, сдержанно, активизируя при этом процесс деления клеток фикобионта. Сторонники мутуалистического симбиоза компонентов лишайников считают, что фикобионт и микобионт находятся в гармоническом сожительстве, приносящем им взаимную выводу: водоросль доставляет грибу органические соединения, а гриб водоросли — воду и минеральные вещества. Но подобные взаимоотношения вряд ли могут существовать в природе, где постоянно происходит борьба между живыми организмами. О том, - что между компонентами лишайника складываются сложные взаимоотношения, может свидетельствовать медленный рост лишайников. Благодаря незначительному накоплению органического в щества ежегодный прирост слоевищ невелик. Так, накипные лиша ники нарастают в среднем на 1-—8 мм в год, листоватые и кустистые- от 0, 1 до 3, 5 см в год, виды кладоний — на 2—5 см. Таким образом, проблема взаимоотношений микобионта и фик> бионта в настоящее время не вполне ясна. Вероятнее всего, взаимоо ■ ношения микобионта и фикобионта в отдельных случаях неоднозначн и зависят от многих факторов. Сложные взаимотношения между микобионтом и фикобионтом лишайнике могли возникнуть в результате длительной эволюции. Пр этом возможны два пути формирования лишайникового симбиоза. Пе] вый путь — от безразличных взаимоотношений между симбионтам; когда гриб питается только за счет слизистых выделений водоросл] к дальнейшему усложнению их, когда к чисто сапротрофному тип питания примешивается элемент паразитизма, при котором микобиоь усваивает некоторые продукты метаболизма живых клеток, и, наконе! к случаям непосредственного паразитизма, когда гриб образует гаустс рии, внедряющиеся в клетки водоросли. Поскольку гаустории образу ются не-всегда, трудно судить о том, имеется ли здесь угнетение одног симбионта", другим, или длительное сосуществование их может продол житься неопределенно долго. Другой путь формирования лишайнике вот симбиоза представляют случаи резкого паразитизма гриба н водоросли, когда гифы гриба захватывают колонии водоросли и микс бионт при помощи гаусторий питается содержимым клетки. В онтоп незе лишайника развивается симбиоз обоих компонентов, при которо! преобладает то один, то другой симбионт в зависимости от условш внешней среды и складывающихся взаимоотношений фикобионта микобионта. ч Компоненты лишайников Микобионты лишайников относятся в основном к сумчатым грибам - пиреномииетам и дискомицетам. Лишь у некоторых тропических и суб тропических видов гриб принадлежит к базидиомицетам. Микобионт! небольшого количества лишайников — грибы с неклеточным мицелие! и несовершенные грибы.. Микобионт представлен тонкими (около 3—10 мкм в диаметре простыми или ветвящимися тифами чс двухслойной оболочкой, харак те'ризующимися верхушечным ростом. Гифы разделены поперечным: перегородками на клетки, протопласты которых соединены тончайшим! цитоплазматическими тяжами (плазмодесмами) через маленькое отвер стие (перфорацию) в перегородке. Некоторые гифы имеют толстук оболочку, способную разбухать, впитывая воду и удерживая ее. В обо лочках гиф откладываются пигменты, придающие лишайникам свое образную окраску. Имеются специфические для лишайников жировьк клетки (или жировые гифы), в которых жир содержится в виде неболь ших капель. Переплетаясь, гифы микобионта образуют плектенхим) (ложную ткань), составляющую основу разнообразно дифференциро ванных слоевищ лишайников. Фикобионты большинства лишайников относятся к зеленым, а у меньшинства — к синезеленым водорослям. Водоросли лишайников по сравнению со свободноживущими значительно изменены. Так, в слоевищах лишайников у водорослей не развиваются слизистые обвертки, уменьшается количество отлагающихся запасных питательных веществ, размеры клеток увеличиваются; колониальные и нитчатые водоросли лишайников часто распадаются на отдельные клетки. Зеленые водоросли лишайников" представлены родами: требук- сия — ТгеЬоихьа, палмелла -— Ра1те11а, глеоцистис — С1оеосузИз, кок- комнцес — Сосеотусез и другими в виде одноклеточных микроскопически мелких шариков. Из нитчатых зеленых водорослей наиболее широко распространены Тгеп1ероНИа, принимающая зеленую окраску в слоевище лишайника, С1айорНога. Зеленые водоросли в слоевище лишайника размножаются простым делением или образуют автоспоры внутри материнской клетки. Из синезеленых водорослей часто встречаются N03(00, АпаЬаепа, С1оеосарза, СНгоососсиз, ЗИдопета, калотрикс — СаШНпх, дихо- трикс — Бьско^пх, хиелл'а — НуеНа и др. В лишайниках у них не образуются гормогонии и споры. Носток в слоевище лишайника, распадается на цепочки клеток, иногда располагающиеся группами, я в неслизистых лишайниках — даже на отдельные клетки.? Из желтозеленых (разножгутиковых) водорослей в качестве фикобионта известны представители рода гетерококку'с — Не1егосоЬсиз. До 90 % лишайников содержат водоросли родов ТгеЬоихьа, Тгеп- ^ероНиа или N03(00. Всего в лишайниках встречаются виды 26—28 родов водорослей. Будучи выделенными из лишайников в чистую культуру, водоросли могут вновь образовывать зооспоры и даже гаметы (зеленые водоросли), восстанавливать морфологию свободноживущих форм, гормогонии и споры (синезеленые). В лишайниках имеются фикобионты, неизвестные в свободном состоянии (ТгеЪоихьа, кокко- микса — Соссотуха, лобококкус — ЬоЬососсиз, диплококкус — О1р1о/Со& * сиз и др.). Морфология и анатомическое ^ строение лишайников
Форма' и величина лишайников от нескольких миллиметров до десятков сантиметров. По форме различают три основных морфологических типа (жизненных форм) слоевищ лишайников: накинный (корковый), листоватый и кустистый. Между ними существуют переходные формы. Иногда выделяют лепрозные (чешуйчатые) и филаментозные (нитевидные) типы слоевищ. Наиболее просто организованы накипные или корковые слоевища, имеющие вид порошковатых, зернистых, бугорчатых, гладких налетов или корочек, плотно срастающихся с субстратом и не отделяющихся от него без значительных повреждений. У некоторых лишайников слоевище так врастает в субстрат, что присутствие лишайника обнаруживается по изменению окраски субстрата и по плодовым телам микоби- онта, заметным в виде темноокрашенных точек или полос (рис. 336). Более высокоорганизованные лишайники имеют листоватое слоевище в форме дорзовентральных пластинок, распростертых по субстрату и срастающихся с ним при помощи пучков грибных гиф, называемых ризинами. У немногих видов слоевище прикреплено к субстрату в одном месте при помощи слоевищного выроста — гомфа, образованного грибными гифами.* На субстрате листоватые лишайники имеют вид чешуек, розеток или довольно крупных, обычно разрезанных на лопасти пластинок (рис. 337). Еще выше организованный тип слоевища— кустистое, имеющее форму ветвящихся лент или более толстых, часто разрезанных на лопасти разветвленных стволиков, срастающихся с субстратом только основанием. Кустистые лишайники растут или вертикально вверх от субстрата, или вбок, или свисают вниз в виде более или менее длинных прядей (рис. 338). Промежуточная форма между накипными и листоватыми такая, у которой слоевище в центре накипное, а по краям листоватое. Между листоватыми и кустистыми также имеются переходные формы (например, виды эверния — Еьегша).
Анатомически различают два типа слоевищ лишайников: гомеомерный[37] и гетеро- мерный[38]. В более примитивных — гомео- мерных — клетки фикобионта распределены равномерно в толще слоевища и в слизи, выделяемой ими, по всем направлениям проходят грибные гифы (рис. 339). Примером могут служить виды коллема — СоИета, нередко встречающиеся на юге на скалах. В сухом состоянии они имеют вид черных ломких корочек или морщинистых подушечек, которые при увлажнении увеличиваются в размерах от разбухания слизи, внутри которой равномерно распределены микобионт и фикобионт. У видов рода лепто- Рис. 339. Вертикальный разрез гомеомер- Рис. 340. Вертикальный разрез гетеромерного слоевища ного слоевища: I — верхняя и нижняя кора. 2 — водоросли (Зона водорослей, альгальная зона, гоиидиальный слой), 3 — сердцевина, 4 — ризины гиум — Ьер1од'шт наряду с беспорядочным расположением компонентов лишайника заметны нижняя и верхняя кора, образованная одним слоем тесно соединенных грибных гиф. Большая дифференцировка слоевища наблюдается у гетеромер- ных лишайников. На поперечном срезе гетеромерного слоевища, например лишайников ксантория настенная — Хап(Нопа рапеИпа, гипо- гимния вздутая — Нуродутта ркузойез, различимы несколько слоев (рис. 340). Сверху слоевище покрыто верхней корой, сформированной плотным сплетением грибных гиф. Это плектенхима. Внутрь слоевища от плектенхимы гифы лежат рыхло и между ними расположены клетки фнкобионта, образующие зону водорослей (альгальная зона, гонидиальный слой). Далее внутри находится сердцевина из рыхло расположенных грибных гиф с большими пустотами, заполненными воздухом. Снизу слоевище покрыто нижней корой, строение которой сходно со строением верхней коры. Из сердцевины через нижнюю кору часто проходят грибные гифы — ризины, при помощи которых лишайники прикрепляются к субстрату. У кустистых лишайников, например у родов уснея — 1}$пеа, алектория — А1ес1опа, имеющих гетеромерно- радиальное строение, на поперечном разрезе видны кора, зона водорослей, сердцевина (рис. 341). У накипных лишайников нижней коры нет, так как они срастаются с субстратом сердцевиной. Внутри каждого из трех основных морфологических типов лишайников могут встречаться как анатомически высокоорганизованные слоевища, так и примитивные. Размножение лишайников У лишайников существуют три типа размножения: вегетативное, бесполое и половое. Размножается либо собственно лишайник, либо мико- бионт. Наиболее часто наблюдается вегетативное размножение, основанное на способности слоевища лишайника регенерировать из отдельных его участков. Оно осуществляется путем фрагментации (отделение участков) слоевища или с помощью специальных образований — соредий, изидий и л о- бул. Фрагментация происходит механически. Хрупкие в сухую погоду лишайники легко ломаются от прикосновения проходящих животных или людей, ими же или ветром переносятся на различные расстояния. Отдельные участки лишайника, попав в соответствующие условия, развиваются в новое слоевище. Соредии — мельчайшие образования, состоящие из одной или нескольких клеток водоросли и окруженные грибными гифами. Они образуются в зоне водорослей. У одних лишайников соредии развиваются вследствие усиленного размножения в слоевище водоросли и оплете- ния ее клеток гифами гриба, у других— усиленно растут гифы и опутывают клетки водоросли. Под давлением образующихся соредий кора прорывается и они выходят наружу в виде порошащегося налета. Обособленные скопления соредий называются соралями (рис. 342). Со- рали обычно имеют форму округлых или продолговатых пятен, разбросанных по поверхности таллома, или каймы по его краю. Если соредии, разносимые ветром, каплями дождя, попадают в условия, благоприятствующие развитию, то возникает слоевище лишайника. В тех случаях, когда в силу неблагоприятных условий соредии не развиваются в новое слоевище, они обильно размножаются, образуя порошковатые пятна, так называемые ленрозные формы. Соредии имеются почти у всех листоватых и кустистых лишайников.
У некоторых лишайников образуются изидии — бугорчатые па- А. Б
Рис. 342. Строение соралей. А—сораль; Б — отдельные соредии лочковидные выросты на верхней поверхности слоевища, состоящие из фикобионта и микобионта (рис. 343). От соралей изидии отличаются тем, что они покрыты корой. Отламываясь, изидии разносятся дождем или ветром и в благоприятных условиях развивают новое слоевище. Являясь выростами слоевища, изидии увеличивают ассимиляционную поверхность, способствуя повышению фотосинтетической активности лишайника за счет фикобионта изидии. Л обул и имеют вид маленьких чешуек, расположенных вертикально на поверхности слоевища или.по его краям. Они напоминают изидии (рис. 344). При бесполом размножении микобионта в пикнидиях образуются пикноконидии, или с т и л о с п о р ы (рис. 345). Пикноконидии лй- Рис. 345. Пикнндии (продольный разрез), А — пикнидии; Б — окончания гиф с пикноконидиями шайников очень разнообразны по форме и величине. Несмотря на то что пикнидии обычно относят к органам бесполого размножения, некоторые лихенологи принимают их за мужские половые органы лишайника, а пикноконидии — за мужские половые элементы. Половой процесс у сумчатых лишайников изучен недостаточно, хотя в общих чертах он аналогичен половому процессу свободноживу- щих грибов. До сих пор у исследователей о половом процессе лишайников нет единого мнения. Одни авторы считают, что у них пикноконидии оплодотворяют женский половой орган — архикарп, состоящий из двух частей: аскогона — в виде спирально закрученной гифы и три'-, хогины — в виде тонкой гифы, отходящей вверх от аскогона (рис. 346). Другие придерживаются мнения, что у лишайников оплодотворение редуцировано: в разрастающемся аскогоне ядра попарно группируются в дикарионы, которые переходят в клетки аскогенныХ гиф, вырастающих из аскогона. Из этих двухъядерных клеток развиваются молодые сумки. По созревании сумок ядра дикариона сливаются в диплоидное ядро (кариогамия). Затем следует редукционное деление (мейоз) ядра с образованием гаплоидных ядер, вокруг которых формируются сумко- споры. Цитологические исследования показали, что в гифах микобионта наблюдается чередование ядерных фаз: гаплоидная фаза (от редукционного деления ядра до объединения ядер в дикарионы), дикариофаза (от группировки ядер в дикарионы до кариогамии), диплофаза (от кариогамии до редукционного деления). У базидиальных лишайников половой процесс еще менее изучен, но имеются данные о том, что вместо оплодотворения осуществляется апогамный процесс. Плодоношения сумчатых лишайников многолетние, а у базидиальных — однолетние или малолетние. Развитие плодовых тел у сумчатых -лишайников протекает двумя путями и на основе их онтогенеза сумчатые лишайники разделяют на две группы: асколоку- лярные и аскогимениальные. У. а с к о л о- кулярных лишайников развитие плодовых тел начинается с возникновения тесного переплетения гиф—.стром, где закладываются архикарпы. Затем в строме появляются особые полости— локулы, в которых формируются сумки со спорами. Кроме сумок в локул ах имеются парафизоиды — остатки стромы в виде вертикально расположенных гиф, срастающихся своими вершинами. У а с к о- < тимениальных лишайников архикарпы закладываются в. верхней части сердцевинного слоя. В плодовых телах формируется настоящий, гимениальный слой из вертикально стоящих сумок со спорами и пара- физ — бесплодных вытянутых гиф, не срастающихся вершинами. У сумчатых лишайников имеются перитеции или апотеции.
Перитеции более или менее погружены в слоевище лишайника и их устьица заметны в виде темных точек. Типичный гимениальный слой, состоящий из сумок и парафиз, располагается на внутренней, чашевидно вогнутой поверхности перитеция и заметен на ранних стадиях развития плодового тела. Впоследствии парафизы исчезают, ослизняясь и растворяясь. Под гимением залагается гипотеций. В верхней части перитеция развиваются перифизы — нежные, нитевидные выросты, защищающие выводное отверстие. Обычно перитеции окружены темного цвета обверткой, состоящей из трех слоев: внутреннего—•
эксципула, среднего — хламиса и наружного — покрышки (рис. 347). Внутренняя полость перитеция заполнена слизью. Апотеции находятся на поверхности слоевища и видны в виде небольших чашечек, блюдечек, бугорков, дисков. Апотеции в виде вытянутых линий называют гастеротециями. В зависимости от строения различают три основных типа апоте- циев: леканоровый, лецидеевый, биаторовый (рис. 348). Эти названия произошли от латинских наименований родов лишайников: леканора — Ьесапога, лецидея — ЬесШеа, биатора — В1а1ога. На разрезе всех типов апотециев заметны следующие слои. Эпитеций — верхняя часть, плодоносящего слоя, где концентрируются" верхуШки " парафиз. Он прикрывает те ц и й, или гнмениальный слой, состоящий из сумок и парафиз. Сумки обычно находятся на разных стадиях развития. Под те- цием развивается ги потец и й,.или субгимениальный слой, где закладываются сумки. У леканоровых апотециев имеется еще слоевищный край, образованный грибными гифами и клетками фикобионта, вследствие чего край апотеция по цвету отличается от диска. Здесь водоросли присутствуют и в слоевище под апотецием. У л е ц и д е е в ы х апотециев развивается собственный край, образованный только гифами гриба. Цвет диска и края лецидеевых апотециев одинаков, обычно черный, слоевищный край отсутствует. Фикобионт отсутствует и в нем, и под апотецием. По строению к лецидеевым апотециям близки биаторовые, отличающиеся более мягкой консистенцией и яркой окраской. Строение апотециев наследственно постоянно и систематически важно. Лецидее- вые и биаторовые апотеции, возможно, более примитивны, чем лекано- ровые, у которых благодаря присутствию водорослей в слоевищном < рае обеспечивается лучшее питание гимения. Сумки у большинства лишайников унитуникатные, имеют одну )болочку, а освобождение спор происходит с помощью верхушечного [апикального) аппарата. У некоторых родов сумки битуникатные, с шумя оболочками. Из таких сумок споры выбрасываются через раз В сумках образуются аскоспоры. Они разнообразны по величине и форме, специфичны для отдельных родов лишайников (рис. 349). Размеры их ^ ^^ колеблются от одного до 300 мкм, наи- 0 1 О: ЩЦ? более обычный размер 10—20 мкм. По п в^й/ форме аскоспоры округлые, веретено- видные, палочковидные, яйцевидные, нитевидные и т. д. Они бывают одно-, дву- и многоклеточные, имеющие только продольные или продольно-поперечные перегородки (муральные). Только у лишайников встречаются биполярные споры — двухклеточные споры с утолщенной перегородкой, разделяющей обе клетки и занимающей •большую часть споры. Толстая перегородка пронизана узким, иногда слабозаметным канальцем. По созревании сумок аскоспоры выбрасываются наружу. Немногие роды — эндокарпон (Епйосагроп) и др.— содержат гй- мениальные водоросли, т. е. фикобионт, попадающий в закладывающиеся плодовые тела и размножающийся в них. Созревшие" споры выбрасываются вместе с фикобионтом, используемым прорастающими спорами грибов для формирования слоевища лишайника. Выбрасываемые из апотециев и перитециев споры лишайникового гриба прорастают в гифы, которые, ветвясь и переплетаясь, развивают зачаточные слоевища, называемые прототалломом. Для дальнейшего формирования настоящего лишайника необходим контакт прото- таллома с определенными водорослями и соответствующие экологические условия. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 618; Нарушение авторского права страницы
 Рис. 336. ОгарЫв вспр1а. Накипное слоевище с гастеро- тециями
Рис. 336. ОгарЫв вспр1а. Накипное слоевище с гастеро- тециями
 Рис. 337. Нуро/уутта ркуво- скз. Листоватое слоевище
Рис. 337. Нуро/уутта ркуво- скз. Листоватое слоевище
 Рис. 338. С1а< Ипа вШ1апв (С1а< 1оша а1рез1г1$). Кустистое слоевище
Рис. 338. С1а< Ипа вШ1апв (С1а< 1оша а1рез1г1$). Кустистое слоевище
 Рис. 341. Продольный (А) и поперечный (Б) разрезы гетеромерно-радиаль-
ного слоевища 11апеа:
1 — коровы й слой, 2 —зона водорослей - (альгальная зона), 3 — сердцевина
Рис. 341. Продольный (А) и поперечный (Б) разрезы гетеромерно-радиаль-
ного слоевища 11апеа:
1 — коровы й слой, 2 —зона водорослей - (альгальная зона), 3 — сердцевина
 Рис. 344. Лооули (/) на апотеции РеШдега сашпа
Рис. 344. Лооули (/) на апотеции РеШдега сашпа
 Рис. 346. СоИета (продольный разрез). Виден многоклеточный архикарп, состоящий из аеко- гона (/) и трихогины (2) -
Рис. 346. СоИета (продольный разрез). Виден многоклеточный архикарп, состоящий из аеко- гона (/) и трихогины (2) -
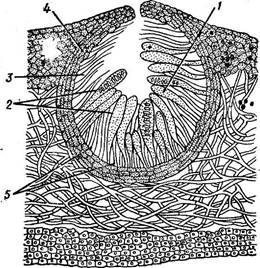 Рис. 347. Перитеций (продольный разрез):
! — гимениальный слой, 2 — сумЛи, 3 — пернфизы, 4 — эксципул, 5 — гнпотеций
Рис. 347. Перитеций (продольный разрез):
! — гимениальный слой, 2 — сумЛи, 3 — пернфизы, 4 — эксципул, 5 — гнпотеций
 Рис. 348. Продольный разрез через апотеции разных типов. Л — биаторовый, Б— лецидеевый; В — леканоровый:
1 — зона водорослей (альгальная зона); 2 — гипотеций, 3 — гнмениальный слой, 4 — слоевищный край, 5 — эпитеций, 6 — эксципул
Рис. 348. Продольный разрез через апотеции разных типов. Л — биаторовый, Б— лецидеевый; В — леканоровый:
1 — зона водорослей (альгальная зона); 2 — гипотеций, 3 — гнмениальный слой, 4 — слоевищный край, 5 — эпитеций, 6 — эксципул