|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ПОДКЛАСС ЭУ АСКОМИЦЕТЫ — Е11А5СОМУСЕТШАЕ
У эуаскомицетов сумки образуются в настоящих плодовых телах, развивающихся по аскогимениальному типу (см. с. 338). Лишь у немногих примитивных представителей этого подкласса сумки формируются группами или пучками на мицелии и не окружены перидием или перидий плодового тела представляет рыхлое сплетение гиф, сквозь которое просвечивают сумки.
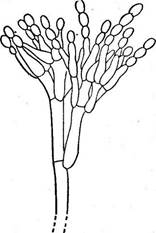
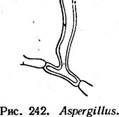
Различают три типа настоящих плодовых тел: ' клейстотеций, перитеций и апотеций. Клейстотеций — округлое, полностью замкнутое плодовое тело, содержащее только сумки. Стерильные элементы — парафизы — в них всегда отсутствуют. Сумки в клейстотециях располагаются неупорядоченно во внутренней плектенхиме (например, у представителей порядка эвроциевые — Еигоиа1ез, рис. 241, А) или образуют в них правильный пучок или слой (у представителей порядка мучнисторосяные — Егуз1рЬа1ез, рис. 241, Б). Созревшие аскоспоры освобождаются из клей- стотеция после разрушения перидия или его разрыва под давлением набухающих сумок. У грибов с прототуникатными сумками аскоспоры из клейстотециев всегда освобождаются пассивно, а для мучнисторо- сяных, образующих в клейстотециях унитуникатные сумки, характерно активное выбрасывание аскоспор. Перитеции — полузамкнутые плодовые тела, большей частью округлые или кувшиновидные, с узким отверстием на вершине. Со дна перитеция, а иногда и от его боковых стенок пучком или слоем поднимаются сумки обычно цилиндрической или булавовидной формы. Между сумками развиваются стерильные нитевидные элементы — парафизы. Настоящие парафизы врастают в полость перитеция между сумками от его основания. Они свободны на концах, простые или ветвящиеся. У представителей порядка гипокрейные —- Нуросгеа1ез образуются парафизы другого типа — апикальные. Это стерильные гифы, врастающие в полость перитеция сверху (с «потолка») и соединяющиеся с его основанием. В этом случае сумки развиваются между апикальными парафизами. Кроме парафиз в перитеции имеются обычно перифизы — нитевидные короткие гифы, расположенные в носике перитеция и направленные к выходу из него (рис. 241, В). Сумки, парафизы и внутренние ткани перитеция называют его центром. В современной систематике эуаекомицетов развитие центра перитеция — важный признак, используемый для разграничения порядков. Аскоспоры из перитециев обычно выбрасываются активно. Упорядоченное расположение сумок в перитеции позволяет им поочередно,, удлиняясь за счет роста или растяжения оболочки, достигать отверстия перитеция и выбрасывать аскоспоры. Аскоспоры выбрасываются из сумки или одновременно, или поочередно, как это наблюдается, например, у видов из рода кордицепс — Согйусерв. У диапортовых — 01арог1Ьа1ез, перитеции которых часто имеют очень длинные носики, сумки образуются на короткой, быстро лизирующейся ножке, и в зрелости свободно лежат в полости перитеция. При набухавши сумок в перитеции создается давление, в результате которого они поочередно выталкиваются к отверстию перитеция и выбрасывают аскоспоры. Лишь у немногих грибов в перитециях развиваются прототуникатные сумки. Их оболочки быстро лизируются, и зрелый перитеций содержит массу аскоспор, погруженных в слизь. При ее набухании споровая масса выдавливается через отверстие перитеция и образует на его вершине слизистую каплю или нить. Апотеции — широко открытые при созревании плодовые тела, обычно блюдцевидные или чашевидные. На их верхней стороне располагается слой сумок и парафиз, называемый гимением. Под гимением находится тонкий слой переплетающихся гиф — гипотеций, или субги- мений. Мясистая стерильная часть апотеция — эксципул — обычно состоит из двух частей: оболочки апотеция, или внешнего эксципула, и его мякоти — внутреннего, или медуллярного, эксципула (рис. 241, Г). Иногда мякоть в апотеции может отсутствовать. У некоторых групп эуаскомицетов строение апотециев отличается от типичного. Они могут иметь булавовидную форму или расчленяться на шляпку и ножку, как, например, у сморчков и строчков. У представителей порядка трюфелевые — ТиЬега1ез апотеции вторично замкнутые в связи с обитанием под землей. Аскоспоры из апотеция выбрасываются активно. Расположение сумок в виде Широко открытого гимения дает возможность одновременного освобождения аскоспор из многих сумок в виде «взрыва» или «залпа». Прикосновение к крупным апотециям пецицевых вызывает появление над ними легкого облачка выброшенных аскоспор и даже звук характерного щелчка. Лишь у немногих аскомицетов, образующих апотеции, аскоспоры освобождаются из сумок последовательно (например, у видов из рода геоглоссум — Сеоц1о8$ит). Аскоспоры выбрасываются из апотециев на расстояние 0, 5—20 см, а у некоторых видов — до 60 см. Апотеций — наиболее совершенный тип плодового тела аскомицетов, обеспечивающий максимальную продукцию аскоспор и их активное выбрасывание на большое расстояние. У многих эуаскомицетов плодовые тела образуются не на мицелии, а на стромах различного размера, окраски и консистенции, состоящих из сплетения гиф. Однако и в этом случае каждое плодовое тело имеет •собственный перидий, хорошо различимый хотя бы на ранних стадиях развития. Система эуаскомицетов основана на строении плодовых тел, типе развития центра перитеция (у пиреномицетов), строении сумок, наличии и характере стромы. В современной системе эуаскомицеты для удобства часто подразделяют на группы порядков соответственно типу плодовых тел и строению сумок, обычно не присваивая этим группам ранга таксонов. К подклассу эуаскомицеты относят сейчас около двадцати порядков. Ниже приводятся наиболее важные из них. Группа порядков плектомицеты. К этой группе относят эуаскомицеты с клейстотециями, реже с перитециями, в которых беспорядочно располагаются прототуникатные сумки. Аскоспоры у этой группы всегда освобождаются пассивно. Основные порядки этой группы следующие: Порядок эвроциевые — ЕигоИа1ез. КлейстотециИ мелкие, развивающиеся на мицелии. Порядок микроасковые — М1Сгоазса1ез. Перитеции содержат беспорядочно расположенные сумки с быстро лизирующимися оболочками. Группа порядков пиреномицеты. Эта группа объединяет эуаскомицеты с перитециями, реже — с клейстотециями, в которых пучком или слоем располагаются унитуникатные сумки. Аскоспоры освобождаются активно. Основные порядки этой группы следующие. Порядок мучнисторосяные, или эризифовы е, — ЕгузЬ- рЬа1ез. Клейстотеции образуются на мицелии. Порядок сферейные — 5рЬаепа1ез. Перитеции темноокра- шенные или черные, обычно кожистой, твердой или углистой консистенции, расположенные на мицелии или в стромах. Порядок диапортовые — 01арог1На1ез. Перитеции, как у предыдущего порядка. Отличается от него сумками с быстро растворяющейся ножкой, в зрелости свободно расположенными в слизи в полости перитеция, а также характером развития центра перитеция. Порядок гипокрейные — Нуросгеа1ез. Перитеции ярко- или светлоокрашенные, мясистой или мягкой консистенции, образующиеся на мицелии или стромах такого же характера, как и перитеции. В перитециях имеются апикальные парафизы, а настоящие парафизы отсутствуют. Порядок спорыньевые, или к л а в и ц е п с о в ы е, — С1ау1С1- рйа1е5. Перитеций и стромы также светло- или яркоокрашенные. Сумки с утолщенной на вершине оболочкой и апикальной порой. Аскоспоры нитевидные, многоклеточные, в зрелости часто распадающиеся на отдельные клетки. От предыдущего порядка отличается также по характеру развития центра перитеция, в частности отсутствием апикальных парафиз. Группа порядков дискомицеты. Плодовые тела у всех представителей этой группы — апотеции. Аскоспоры освобождаются активно. Исключение составляют трюфелевые — ТиЬега1ез — подземные грибы, у которых аскоспоры освобождаются после разрушения перидия плодового тела. Группа включает следующие основные порядки. Порядок гелоциевые—Не1о11а1ез. Сумки иноперкулятные, открывающиеся на вершине трещиной или разрывом (см. рис. 257, А). Апотеции большей частью типичного строения. Порядок фацидиевые—РЬаасИа1е8. Апотеции образуются в стромах и долго прикрыты сплетением мицелия, раскрываются вследствие его разрыва лопастями или щелью. Порядок пецицевые — Рег12а1ез. Сумки оперкулятные, вскрываются на вершине крышечкой (см. рис. 257, Б). Апотеции типичного строения, а также расчлененные на шляпку и ножку. Порядок трюфелевые — ТиЬега1ез. Плодовые тела крупные, клубневидные, подземные, по онтогенезу представляют собой апотеции. Особое положение среди аскомицетов занимает большой порядок лабульбениевые — ЬаЪои1Ъеша1ез. Это группа высокоспециализированных облигатных паразитов насекомых, отличающаяся по морфологии от остальных эуаскомицетов. Лабульбениевые не образуют мицелия, их вегетативное тело представляет рецептакул тканевого строения. На нем развиваются микроскопические перитеции с про- тотуникатными сумками, расположенными пучком. Формально этот порядок может быть включен в группу пиреномнцетов. Некоторые микологи выделяют его в самостоятельный подкласс класса аско- мицеты. ПЛЕКТОМИЦЕТЫ Плодовые тела — клейстотеции, реже перитеции с беспорядочно расположенными прототуникатными- сумками. Освобождение аскоспор всегда пассивное. Порядок эвроциевые — ЕигоНа1е8 X этому порядку относятся эуаскомицеты, образующие клейстотеции с беспорядочно расположенными сумками непосредственно на мицелии. Лишь у немногих его представителей клейстотеции развиваются в стромах, напоминающих склероции. У большинства эвроциевых клейстотеции мелкие, не превышающие 1—2 мм в диаметре. Исключение составляют только оленьи трюфели (род элафомицес — Е1аркотусев), подземные клейстотеции которых достигают нескольких сантиметров в диаметре. Перидий клейстотециев имеет разнообразное строение: от рыхлого паутинистого сплетения гиф, мало отличающихся от вегетативных, до плотного псевдопаренхиматического. Сумки у эвроциевых прототуникатные, с быстро разрушающейся оболочкой, шаровидные или эллипсоидальные, с двумя — восемью аскоспорами. Аскоспоры всегда одноклеточные, бесцветные или окрашенные. Они освобождаются пассивно, после разрушения оболочки сумки и перидия клейстотеция. В распространении большинства эвроциевых существенную роль играет бесполое размножение конидиями. Лишь немногие представители порядка имеют в цикле развития только сумчатую стадию. К порядку эвроциевые относится несколько сотен видов. Большинство из них сапротрофы. Они обитают в почвах различных климатических областей, а также на разнообразных субстратах растительного, реже животного происхождения, на которых они образуют плесени. Некоторые эвроциевые термофилы, развивающиеся на разнообразных саморазогревающихся субстратах (например, в компосте, влажном сене и др.) при температуре 30—60°С. Отдельные виды вызывают заболевания теплокровных животных и растений. Некоторые представители этой группы, поселяясь на пищевых продуктах, промышленных материалах и изделиях, вызывают их плесневение и порчу. В благоприятных для развития условиях, особенно в тропиках, они за короткий срок могут вызывать разрушение тканей, кожи, различных синтетических материалов (например, электроизоляции), ускорять процессы коррозии металлов, повреждать приборы, оптику и многие другие изделия. Среди эвроциевых есть кератинофильные грибы, •образующие " йератинолитические ферменты и развивающиеся на субстратах, содержащих нерастворимый фибриллярный белок кератин, — перьях, рогах, копытах. Известны эвроциевые, патогенные для человека и животных и не: редко вызывающие тяжелые заболевания — дерматомикозьги глубокие микозы. Среди болезней особенно опасен гистоплазмоз, возбудителем которого является ЕттопвьеИа сарвиШа. Многие эвроциевые и родственные им несовершенные грибы из родов аспергилл — АзрегцШив, пеницилл — РетсИИит, акремониум — Асгетотит и других образуют- антибиотики, ферменты, органические кислоты и используются для их промышленного получения. Так, одним из основных продуцентов широко используемого в медицинской практике антибиотика цефалоспорина С является представитель порядка эвроциевые эмерицеллопсис почвенный — Етег1се11ор818 1егпсо1а. На основе этого антибиотика в последние годы были получены полусинтетические производные, которые.не только превосходят исходный препарат по активности, но и действуют на те группы организмов, которые устойчивы к действию пенициллина, — грамотрицатель- ные бактерии и устойчивые к пенициллину стафилококки. Полусинтетические цефалоспорины — цефалотин и цефалоридин — выпускаются про-' мышленностью и используются в медицинской практике. Одну из наиболее важных групп эвроциевых составляют грибы, конидиальные стадии которых относятся к родам РетсИИит и Азрег- цШиз. Эти два рода объединяют многочисленные виды грибов, повсеместно распространенных в почвах от Арктики до тропиков, а также на субстратах растительного происхождения. У многих из них известны только конидиальные стадии, такие виды относят к несовершенным грибаЭто грибы, обычно обитающие в почве, где они активно разрушают органические остатки. Многие из них вызывают плесневение пищевых продуктов (черная и зеленая плесени), повреждение изделий из пластмасс, металлов и других материалов. Некоторые аспергиллы, например А. ]ит'ща1из, служат причиной заболеваний человека и животных— так называемых аспергиллезов. Другой вид этого рода — А. Цаоив, часто развивающийся на плодах арахиса и различных кормах, образует афлатоксин— токсин, вызывающий тяжелые поражения печени. Афлатоксин подавляет синтез ДНК и митоз, стимулирует развитие гигантских клеток и обладает канцерогенными свойствами. Некоторые виды пенициллов поражают плоды (яблоки, цитрусовые) в период хранения. Пенициллы и аспергиллы широко используются в микробиологической промышленности — в производстве органических кислот (лимонной, фумаровой, глюконовой и др.), ферментов (протеи- наз, амилаз и др.) и антибиотиков (пенициллина, гризеофульвина, фумагиллина). Из последних особое значение имеет пенициллин, образуемый грибом РетсИИит скгузодепит. Это один из основных антибиотиков, применяемых сейчас в медицине. У ряда видов пенициллов и аспергиллов, по традиции также относимых к несовершенным грибам, известны сумчатые стадии, относящиеся к разным родам эуроциевых. Поскольку, согласно Международному кодексу ботанической номенклатуры, основным наименованием плео- морфных грибов является наименование их совершенной стадии, эти виды должны рассматриваться среди аскомицетов. Для рода аспергилл характерны простые конидиеносцы, вздутЫе на вершине в виде пузыря различной формы. На нем располагаются ^фиалиды, образующие цепочки одноклеточных гконидий (рис. -242). У некоторых аспергиллов фиалиды находятся не на самом пузыре, а на специальных клетках — профиалидах. 23 Заказ 552 Представители рода эвроциум — Еигойит часто встречаются в природе на разлагающихся растительных субстрата_х. В зависимости от условий среды (температуры, влажности)и соответственно преимущественного развития конидиалыюй или сумчатой стадии они обра-, зуют зеленые, желтоватые или красно-желтые плесени. Клейстотеции видов этого рода обычно шаровидные, очень мелкие, желтые, с однослойным перидием, покрыты рыхлой сетью гиф с желтоватыми или красноватыми гранулами. Сумки образуются на аскогенных гифах по способу крючка. Оболочки сумок разрушаются очень рано, и зрелые клейстотеции содержат массу бесцветных или желтоватых линзовидных аскоспор с экваториальной бороздкой. Обычный субстрат для многих видов из этого рода — хранящиеся растительные продукты. Многие из его представителей ксерофилы, развивающиеся в условиях пониженной влажности, неблагоприятных для роста других грибов. Например, эвроциум ползучий (Е. герепз) вызывает плесневе- ние зерна и многих других продуктов при влажности 13—15%. Большое значение имеют эвроциумы и как возбудители биоповреждений промышленных изделий и материалов. Развиваясь при малых значениях влажности и минимуме питания, они вызывают разложение текстиля, целлофана, резины, пластмасс, встречаются даже на стекле оптических приборов, вызывая их порчу. Они ускоряют также процессы коррозии металлов, вероятно, вследствие образования большого количества органических кислот. Виды эвроциумов обычно осмофильны, т. е. могут развиваться на средах с высоким осмотическим давлением, например с повышен- яым содержанием сахара (до 20 % и более). Е. герепз, например, часто встречается на заплесневелых джемах и вареньях, где образует обильные конидии и клейстотеции. Для второго крупного рода эвроциевых с конидиальной стадией аспергилл — эмерицелла (ЕтепсеИа)—характерны шаровидные, довольно крупные клейстотеции, обычно ярко-желтые, окруженные массой крупных толстостенных клеток. Перидий клейстотециев состоит из нескольких слоев переплетающихся гиф. Аскоспоры лйнзовидные, с экваториальными гребнями, красные или фиолетовые. Виды этого рода встречаются повсеместно в почвах, а также на растительных материалах.
Один из наиболее распространенных видов — эмерицелла лежачая (ЕтепсеИа тс! и1апз) образует быстрорастущие колонии, окраска которых варьирует от зеленой до ярко-желтой в зависимости от степени развития конидий и клейстотециев. Клейстотеции желтые, хорошо видны невооруженным глазом. Этот вид обычен на растительных субстратах, встречается в почвах умеренной зоны и субтропиков, иногда развивается в дыхательных путях теплокровных животных. Этот вид широко используется в генетических исследованиях для изучения закономерности образования гетерокарионов, в том числе генетического контроля этого процесса, цитоплазматиче- ской наследственности, парасексуального процесса и других вопросов. У видов рода пеницилл конидиеносцы имеют вид кисточки. Их строение разнообразно. Наиболее простые кисточки состоят из одной мутовки фиалид на вершине кони- диеносца. Более сложная кисточка состоит из метул — более или менее удлиненных клеток, расположенных на вершине кони- диеносца, на каждой из которых в свою очередь располагается мутовка фиалид. Такие кисточки бывают как симметричными, так и асимметричными. Конидиеносцы могут иметь и еще более сложное строение: на его вершине образуются веточки, на них — ме- тулы, а на метулах — мутовки фиалид (рис. 243). Сумчатые стадии известны у немногих видов этого рода. Примером эвроциевых с этой кониди- альной стадией может служить род таларо- мицес—Тсйаготусея, виды которого образуют мелкие шаровидные или неправильной формы клейстотеции с неограниченным ростом. Клейсто- теции обычно имеют ярко-желтую или оранжево-желтую окраску и, появляясь в изобилии, придают характерную желтую окраску колониям грибов этого рода. У широко распространенного в почвах таларомицеса желтого — Т. Цаюи5 в культуре образуется сначала конидиальная стадия — пеницилл червеобразный — РепШШит ьегт1си1а1ит. Затем на гифах развиваются спирально закрученные аскогоны и антеридии. После оплодотворения вырастают аскогенные гифы, а из них цепочками образуются сумки. Этот вид часто вызывает биоповреждения различного оборудования и материалов, особенно в тропиках и субтропиках. Порядок микроасковые — М1" сгоа5са1е5
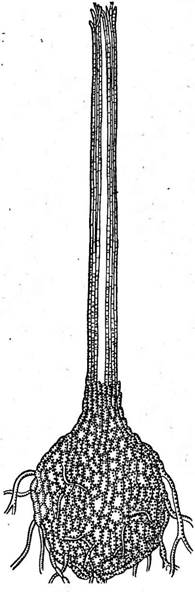
Небольшой порядок микроасковые объединяет грибы с тёмноокрашен- ными перитециями, содержащими беспорядочно расположенные про- тотунйкатные сумки. Перитеции образуются на поверхности субстрата или частично в него погружены. Они имеют шаровидную или грушевидную форму и часто длинный носик, по длине в несколько раз превышающий диаметр перитеция (рис. 244). Парафизы в перитециях отсутствуют, но в их устье образуются перифизы. Оболочки сумок быстро лизируются, и зрелые перитеции содержат массу аскоспор, погру Микроасковые развиваются, как сапротрофы на разнообразных растительных субстратах или как паразиты высших растений. Некоторые из них вызывают серьезные болезни растений, приносящие большой ущерб сельскому и лесному хозяйству, например голландскую болезнь вязов, увядание дуба, гнили различных растений. Известны микроасковые, вызывающие синеву древесины. Наиболее важный род из этого порядка — род цератоцистис (Сега1о- сузНз). К нему относятся такие вредоносные паразиты растений, как цератоцистис вязовый (С. и1гтй)—возбудитель голландской болезни вязов и бахромчатый — С..]1тЬг1а1а, вызывающий черную гниль батата и болезни многих других растений. Интересную группу видов этого рода представляют грибы — возбудители синевы древесины сосны, ели и некоторых лиственных деревьев. Заболонная часть древесины окрашивается в синий или серовато-синий цвет. С. и1пи — один из наиболее известных и важных видов этого рода. Он вызывает голландскую болезнь вяза» распространенную в Европе и Северной Америке и приносящую большой ущерб лесам, парковым и защитным насаждениям. У пораженных этим грибом деревьев внезапно желтеют и увя- " 41 " дают листья и усыхают концы ветвей. Рис. 244. Оега1осу$И&. Перитеций ПРИ сильном поражении все дерево может усохнуть в течение нескольких дней. Эта болезнь впервые появилась в Голландии в начале нашего века, а затем распространилась по всей Европе и была завезена на американский континент.
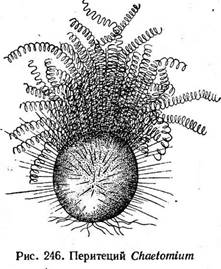
Весной под корой больных деревьев развиваются черные коремии конидиального спороношения этого гриба — графиума вязового— Ога- ПИРЕНОМИЦЕТЫ Плодовые тела перитеции, реже" клейстотеции, в которых пучком или слоем располагаются унитуникатные сумки. Освобождение аскоспор активное. Порядок мучнисторосяные, или эризифовые, — Егу81рЬа1е$ ' Порядок мучнисторосяные объединяет эуаскомицеты, образующие клейстотеции, в которых сумки в' зрелости располагаются правильным пучком или слоем (см. рис. 241, Б). Представители этого порядка — обли- гатные паразиты высших растений, вызывающие у них заболевания, известные под общим названием «мучнистая роса». В культуре на искусственных питательных средах'эти грибы пока не получены. Белый, позднее темнеющий мицелий мучнисторосяных грибов находится обычно на поверхности пораженных органов растений. На нем образуются специальные структуры — апрессории, от которых отходят гаустории, проникающие в клетки эпидермиса растения-хозяина. У некоторых представителей этого порядка гаустории могут внедряться 'в мезофилл листа (например, у видов из рода филлактиния— РНуИасИ- та). У грибов из рода левейюла—ЬеюеШи1а, распространенных в засушливых районах, мицелий развивается внутри тканей растения и частично выходит через устьица на его поверхность. Образование мицелия внутри тканей растения — приспособление гриба к обитанию в условиях низкой влажности. На мицелии мучнисторосяных через несколько дней после заражения развивается конидиальная стадия — прямые неразветвленные конидиеносцы с цепочками конидий (рис. 245, А). У некоторых представителей этого порядка на конидиеносцах образуются одиночные конидии (например, у видов рода ^еVе^^^и^а). В это время пораженные органы растения покрыты мучнистым налетом конидий — отсюда название заболевания'—«мучнистая роса». Конидии распространяются воздушными течениями и заражают новые растения. Мучнисторосяные грибы — ксерофилы. Их конидии могут развиваться и прорастать в сухую погоду при относительной влажности воздуха до 60 %. Сумчатая стадия развивается у мучнисторосяных в конце периода вегетации. На мицелии гриба образуются аско^оны (без трихогины) и антеридии. Содержимое антеридия переходит в аскогон, который делится после этого на ряд клеток, одна из которых содержит дикарион. Эта клетка или непосредственно трансформируется в сумку (у мучни- Рис. 245. Порядок Егу51рЬа1е$. А — конидиеносцы с конидиями; Б—Е — клейстотеции: Б — Егуйрке, В — ЗрНаегоИгеса; Г — М1сго& рНаега; Л — Ь: папи1а; Е—РНу11асИта сторосяных, образующих только одну сумку в клейстотеции), или из нее развиваются аскогенные гифы, а на них — сумки. Одновременно формируется перидий клейстотеция. Наружный и внутренний слои перидия различаются морфологически и функционально. Наружный слой его состоит из толстостенных гиф и является защитным, а внутренний — из тонкостенных, быстро лизирующихся гиф — выполняет питательную функцию. Из наружных слоев перидия развиваются придатки, или аппендиксы, различного строения (рис. 245). Их форма —характерный признак родов мучнисторосяных, так же как и число сумок в клейстотеции. Развитие клейстотециев и сумок — длительный процесс. Сумки созревают только осенью, а у некоторых мучнисторосяных — к весне. Зимующей стадией являются клейстотеции, однако у некоторых грибов из этой группы мицелий может сохраняться в зимующих частях растений. В странах, с теплым и мягким климатом клейстотеции часто совсем не образуются. Аскоспоры освобождаются из сумок активно. Сумки с созревшими аскоспорами набухают и разрывают перидий клейстотеция. Дальнейшее набухание сумок вызывает разрыв их оболочки, и аскоспоры разбрасываются во всех направлениях. Мучнисторосяные паразитируют на растениях из разных семейств, однако большинство из них (около 90%) развивается на двудольных. Среди немногочисленных паразитов на однодольных наибольшее значение имеет возбудитель мучнистой росы злаков — эризифе злаковая (Егу$1рНе цгатШ& ). Этот вид распадается на несколько специа- лизированных форм, приуроченных к отдельным видам или группам близких видов. Он поражает пшеницу, рожь, ячмень и другие злаки. Клейстотеции Е. дгатШз, как и других видов этого рода, содержат несколько сумок и имеют придатки, напоминающие вегетативные гифы (рис. 245, Б). Такие же придатки характерны и для другого рода мучнисторосяных— сферотека (8ркаего1кеса), однако у его представителей в клейстотеции образуется только одна сумка (рис. 245, В). Один из наиболее распространенных видов этого рода — сферотека крыжовника (5. тогз тае)—возбудитель американской мучнистой росы крыжовника. Этот гриб происходит из Северной Америки, откуда он был завезен во многие страны. Он поражает ягоды, стебли и листья крыжовника,, а также смородину. При сильном заражении кусты гибнут. Ягоды покрываются войлочным налетом мицелия гриба, сначала светлым, а затем темнеющим, они не созревают и теряют товарную ценность. Для представителей рода АИсгозркаега характерны жесткие экваториальные придатки, дихотомически ветвящиеся на концах (рис. 245, Г). В клейстотециях образуется по нескольку сумок. Широко распространена микросфера альфитоидес (М. а1ркИоШе5) — возбудитель мучнистой росы дуба, сильно поражающая молодые побеги и поросль, часто вызывая их засыхание. На листьях и стеблях молодых побегов образуется характерный белый порошистый налет конидий, а к осени развиваются клейстотеции. Мучнисторосяные грибы из рода унцинула — С1псти1а имеют слегка вдавленные снизу клейстотеции с многочисленными жесткими придатками, спирально закрученными на концах (рис. 245, Д). К этому роду относится возбудитель пейелицы винограда — унцинула некатор (С/, песа(ог), поражающий листья, ягоды и стебли винограда и приносящий в годы массовых вспышек большие потери урожая. Как и у возбудителя мучнистой росы крыжовника, родина этого гриба — Северная Америка, откуда он был завезен в Европу в середине прошлого века. Наиболее сложно устроенные придатки имеют клейстотеции грибов из рода филлактиния — РкуИасИта. Оуи двух типов: на вершине клейстотеция — в виде тонких, ветвистых гиф, выделяющих слизь, а по экватору клейстотеция — в виде шипов, расширенных у основания в виде луковицы. Оболочка этой расширенной части снизу более тонкая, при высыхании сморщивается, в результате чего экваториальные придатки отгибаются вниз, приподнимая клейстотеций над поверхностью субстрата (рис. 245, Е). Он легко сдувается ветром и, попав на какой-либо субстрат, приклеивается к нему слизью, выделяемой придатками на. вершине клейстотеция. Порядок сферейные — 8рЬаепа1е5 Для порядка сферейные характерны типичные кувшиновидные перитеции с хорошо развитым темным перидием, обычно пленчатым, кожистым или твердым. В перитециях содержатся булавовидные или ци При половом процессе у сферей- ных наблюдается как типичная для аскомицетов гаметангиогамия, так и разнообразные отклонения от типа. Так, у многих антеридии не функционируют или вообще утрачены. В этом случае часто наблюдается сперматизация (например, у видов рода подоспора — Ройозрога, нейроспоры густой — Ыеигозрога сгазза). У большинства сферейных ас- коспоры выбрасываются из перитеция активно: сумки поочередно вытягиваются, высовываются в отверстие перитеция и выбрасывают аскоспоры. После этого оболочка пустой сумки спадается, и ее место занимает следующая сумка. У примитивных представителей порядка активного выбрасывания спор не наблюдается. Примером наиболее примитивных сферейных может служить -род хетомиум — СНае1от1ит. Виды этого рода распространены на растительных остатках и в почве, где они играют-существенную роль в раз- ' ложении растительного опада. Нередко они встречаются и на разнообразных целлюлозосодержащих материалах и изделиях, например бумаге, книгах и др., которые они могут повреждать. Перитеции у представителей этого рода покрыты волосками (рис. 246), форма которых служит здесь видовым диагностическим признаком. Оболочка сумок у них быстро лизируется, и зрелый перитеций содержит массу аскоспор, погруженных в слизь. Их выход из перитеция происходит так же, как у микроасковых: слизь набухает, и аскоспоры выходят из перитеция в длинном слизистом шнуре. У копрофильных грибов, из рода сордария — Богдана перитеции имеют тонкий пленчатый перидий бурого цвета и образуются на мицелии. Парафизы рано разрушаются, и зрелый перитеций содержит только сумки. Аскоспоры у сордарий темноокрашенные, со слизистой обверткой (рис. 247). Наиболее распространенный вид этого рода—- сордария навозная — 5. /1т1со1а — не образует конидиального споро- ношения и размножается только аскоспорами. Носик перитеция у сор- дарии положительно фототропичен. Выброшенные из перитеция аскоспоры благодаря слизистым обверткам прилипают к траве, попадают с ней в кишечный тракт травоядных животных, а затем, попав на навоз, прорастают.
Виды этого рода хорошо растут в культуре на питательных сре-. Рис. 247. 5 огйагьа. Перитеций: А — внешний вид; Б —разрез; В — сумка; Г — аскосиора дах, образуя многочисленные перитеции, поэтому их широко используют для изучения физиологии и генетики аскомицетов. Сордария представляет очень удобный объект для генетических исследований. Аскоспоры у нее крупные и расположены в сумках в один ряд, поэтому их легко можно извлечь оттуда в определенной последовательности при помощи микроманипулятора. Получены мутанты сордарий с желтыми и серыми аскоспорами, используемые для скрещивания и последующего генетического анализа. Особенно широко используют в генетических исследованиях виды близкого к сордарии рода нейроспора — Ыеигозрога. Наиболее популярный объект генетики грибов — нейроспора густая — N. сгазза. Аскоспоры у нее коричневые, с продольной орнаментацией, откуда и происходит название рода (от греч. «нейрос» — жила). Этот вид гетеро- талличен. На мицелии гриба образуются протоперитеции — аскогоны, окруженные стерильными гифами. От аскогона отходят длинные ветвящиеся гифы—так называемые «поисковые гифы», функционирующие как трихогина. Если поисковые гифы встречают конидию или вегетативную гифу совместимого типа, происходит оплодотворение. Из оплодотворенного аскогона развиваются аскогенные гифы, а на них по способу крючка — сумки. Другой вид этого рода — нейроспора ситофила — N. зЫорНИа — благодаря образованию многочисленных, легко распространяющихся конидий и быстрому росту часто вызывает заражение культур в лабораториях. У строматических сферейных из родов гипоксилон — Нуроху1оп и ксилярия — Ху1апа строма обособлена от субстрата, темноокрашен- ная, состоит только из гиф гриба. В нее погружены перитеции с хорошо заметным собственным перидием. Носики перитециев выступают из стромы. У обитающих сапротрофно на древесине видов рода Нуроху1оп стромы имеют полушаровидную или клубневидную форму (рис. 248, А, Б). Представители другого рода — Ху1апа — образуют хорошо дифференцированные стромы, расчлененные на стерильную и фертильную части. Виды этого рода обитают на древесине, реже на других субстра-" тах и особенно распространены в тропиках. Их стромы имеют булавовидную форму или ветвятся в виде оленьих рогов (рис. 248, В). На стромах сначала развивается кони- диальная стадия, а затем перитеции. Порядок диапортовые — Р1арог1На1е$ Перитеции диапортовых внешне сходны с перитециями предыдущего порядка — сферейных. Они имеют плотный кожистый перидий и окрашены в бурый или черный цвет. В носике перитеция находятся перифизы, однако парафизы всегда отсутствуют. Центр перитеция сначала псевдопаренхиматический, но по мере развития сумок эта псевдопаренхима разрушается. Сумки диапортовых имеют короткую ножку, лизирующуюся при созревании, поэтому в зрелом перитеции сумки свободно лежат в его полости, погруженные в слизь. Верхняя часть оболочки сумки утолщена, и споры выходят из сумки через узкий канал. Как и у большинства других аскомицетов, паразитирующих на растениях, у диапортовых конидиальные стадии образуются на живом растении, а перитеции развиваются после гибели растения или его частей на отмерших листьях, коре и ветвях деревьев и других растительных субстратах и обычно погружены в них. У многих представителей порядка перитеции формируются в стромах различного строения. К этому порядку относятся многие возбудители болезней растений, например, гломерёлла опоясанная — С1отегеЫа Ып^иШа — возбудитель горькой гнили плодов яблони и груши, эндотия паразитическая— ЕпйоШа рагазШса, вызывающая рак каштана, возбудители антракноза (сухой гнили) многих растений. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 362; Нарушение авторского права страницы
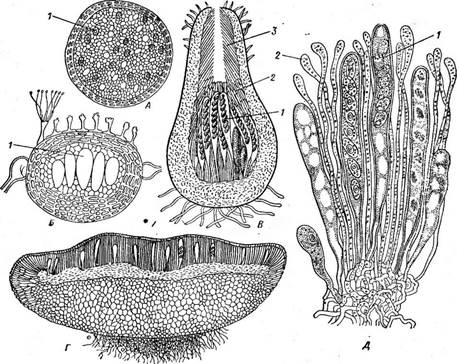 Рис. 241. Типы плодовых тел аскомицетов. А, Б — клейстотеции; В — перитеций;
Г, Л —апотеций:
1 — сумки, 2 — парафизы, 3 —перифизы
Рис. 241. Типы плодовых тел аскомицетов. А, Б — клейстотеции; В — перитеций;
Г, Л —апотеций:
1 — сумки, 2 — парафизы, 3 —перифизы
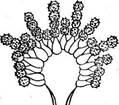
 Строение конидненосца
Строение конидненосца