
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Порядок дрожалковые — Тгеше11а1е8
Порядок объединяет базидиомицеты, имеющие шаровидные или слегка продолговатые четырехкле- то^ные базидии, разделенные продольными взаимно перпендикулярными перегородками на четыре клетки, лежащие рядом. Семейство сиробазидиевые (31ГоЬа- 51сНасеае) включает один род сиробазидиум — 51го- ЬазМшт, характеризующийся базидиями, располагающимися в цепочках. У них очень короткие стеригмы, поэтому базидиоспоры кажутся развивающимися непосредственно на базидии. В семействе дрожалковые (ТгетеНасеае) базидии одиночные, образующиеся на фертильных гифах. Стеригмы отчетливо выражены, развиваются от каждой клетки базидии и выходят на поверхность плодового тела (рис. 315). Базидиоспоры прорастают вторичными спорами или конидиями, впоследствии почкующимися, в результате чего формируются дрожжевидные колонии. Эту стадию развития некоторые авторы по ряду хемотаксономических признаков (состав ка- ротиноидов, содержание ГЦ-оснований и др.), микро- и макроморфо- логических характеристик сближают с аспорогенными дрожжами из рода криптококкус — Сгур1ососсиз, считая их несовершенной стадией
отдельных родов порядка Тгеше11.а1е8. Плодовые тела разнообразны по форме и строению: от тонких, широкораспростертых до крупных, студенистых. Тонкие, широкораспростертые плодовые тела имеются у видов рода эксидиопсис — Ех1- й'юрзьз, развивающихся на гниющей древесине. Эксидиопсис известковый — Е. са1сеа образует крупные плоские плодовые тела серовато-белого цвета, зависящего от кристаллов оксалата кальция, покрывающих гифы гриба и образующихся в процессе обмена веществ. Виды эксидия — Ех1- сНа с крупными мармеладовидными коричневыми или черными плодовыми телами хорошо заметны во влажную погоду осенью и весной на отмерших веточках деревьев и кустарников. В сухую погоду они принимают вид корочек. Представители рода дрожалка — ТгетеИа отличаются от Рис..'315. Разрез через предыдущего яркой (желтой, оранжевой) окрас- плодовое тело ТгетеИа кой плодовых тел И формой спор. У ЕхШа бази- Большинство видов этих родов — сапротрофы на древесине (экси- дия железистая — ЕхШа д1апйи1оза, дрожалка оранжевая— ТгетеНа тезеп1епса), но'средц них есть и паразиты: дрожалки буковая—- Т. ]а^1пеа на буке и гриболюбивая — Т. тусе1орЫ1о1йез — микопаразит на афиллофоровых грибах. г_ — * 7 Представители порядка /дакримицетовы^Г'(Расгуту~се1а1е5) ^имеют нер аз делетпш^баМдии! У дакримицетов базидии цилиндрической формы, на конце виль- чато разветвленные на два отростка, на каждом из которых развивается по одной базидиоспоре. У многих дакримицетов базидиоспоры многоклеточные. Плодовые тела дакримицетов разнообразны: широкораспростертые, булавовидные, кустовидные, студенистые, хрящеватые. Все они сапротрофы на древесных остатках. На опавших ветвях хвойных пород развивается дакримицёс деликвесценс — Пасгутусез йеН- циезсепз в виде мягко-студенистых оранжевых или желтых бугорков и подушечек. Калоцера вискоза— Са1осега V^8С08а имеет роговидные кустисто разветвленные ярко-оранжевые плодовые тела, напоминающие рогатиковые, часто произрастающие на почве во влажных сырых местах (рис.316). Во всех разобранных систематических группах подкласса Не1его- ЪаеийютусеШае заметен параллелизм в форме плодовых тел: представители названных порядков имеют плодовые тела, напоминающие плодовые тела представителей других систематических групп класса Ва51сНотусе1е§. Это свидетельствует о том, что в процессе приспособления к условиям окружающей среды в отдаленных таксонах грибов в результате конвергентной эволюции возникают сходные по форме плодовые тела. В пределах разбираемой группы можно проследить эволюцию плодовых тел от корочек, распростертых по субстрату, до кустовидно разветвленных и даже замкнутых плодовых тел, встречающихся в иных группах- базидиомицетов.
В последнее время гетеробазидиаль- ные грибы не всеми микологами рассматриваются как однородная монофилетиче- ская группа, поэтому имеется тенденция ликвидировать этот подкласс или изменить его объем. При этом в подклассе Не^егоЪазЫютусеМае остается только порядок Тгеше11а1е5. Порядок Аипси1а- па1е5 переносится в следующий подкласс— ТеПоЬазЫютусеийае, где он помещается вместе с головневыми и ржавчинными грибами на основании того, что все они имеют четырехклеточные базидии, а порядок ОасгутусеЫез в ранге семейства переносится в порядок АрЬу1- 1орЬога1е5, так как базидия этих грибов рассматривается как простая (холобази- ( ПОДКЛАСС ТЕЛИОБАЗИДИОМИЦЕТЫ, или склеробазидиомицеты, — те1л0ва8ю! 0мусетюае, зсьековазтюмусетюае Подкласс включает базидиальные грибы, у которых_базидия вырастает из толстостенной покоящейся клетки — телиоспоры, или телейтоспоры. Эти споры выполняют функции стадий, в виде которых гриб переносит неблагоприятные условия, а у большинства видов — зимует. Плодовых тел в подклассе нет, они утрачены вследствие паразитического образа " жизни. Телиоспора " рассматривается также как предшественник базидии — пробазидия. Телиобазидиомицеты разделяют на два порядка: головневые — 1ЬШа& ша1е8 и ржавчинные — 11гес1та1е8. Все грибы, входящие в этот подкласс, — паразиты растений. Порядок головневые —1МПа §ша1е§ Головневые грибы — паразиты многих цветковых растений. В СССР их насчитывается окбло~330 видов. Грибы эти поражают цветки (тычинки, завязи), семена, вызывая их разрушение (рис. 317), а также листья и стебли, иногда корни, на которых появляются характерные вздутия в виде полос или галлов темного цвета. Иногда наблюдается деформация всего растения или отдельных его органов. Разрушенные" репродуктивные органы превращаются в темную __пылящую или мажущую массу — скопление покоящихся спор, называемых телиоспорами или головневыми спорами. Внешне это напоминает обгорелую головешку — отсюда и название порядка. Телиоспоры скапливаются также во вздутиях пораженных вегетативных органов. Мицелий головневых грибов распространяется по межклетникам тканей зараженных растений, процц^ая в клетки при помощи гаусторий. Мицелий двухъядерный (дикарйоти^еский). Нахождение его в тканях до известного времени мало сказывается на внешнем облике растений. Например, при поражении головневым грибом растений овса до времени выколашивания больные растения отличаются от здоровых лишь небольшим отставанием в росте и только после появления метелок можно видеть, что вместо зерен колоски заполнены черной пылящей массой — телиоспорами гриба. У некоторых головневых грибов скопление телиоспор на вегетативных органах становится заметным вскоре после заражения растений, например при заражении кукурузы пузырчатой головней. " Л 1.1 " " " Ь.41 Телиоспоры прорастают четырехклеточной, реже одноклеточной базидией (рис. 318). Дальнейшее развитие гриба (после появления базидиоспор), а также образование двухъядерности у разных видов неодинаково и может быть сведено к трем основным типам. ^ У возбудителя головни проса — ЗрНасеШНеса раш- Ш-тШасеь телиоспоръ1 прорастают четырехклеточной базидией, в которой редукци- онно делится ядро й образуются гаплоидные ядра; каждое из ядер переходит в развивающиеся базидиоспоры. Базидиоспоры или непосредственно копулируют между собой с образованием копуляционных мостиков, или сначала почкуются, а затем копуляция происходит уже между почкующимися клетками. В обоих случаях после коупуляции развивается дикариотический мицелий., На базидии развивается равное количество базидиоспор с разными половыми знаками — « + » Л—-& -—», копулируют базидиоспоры только разных половых знаков. Растения заражаются дикариотичным мицелием. Гриб, прорывает ткани молодых проростков до их выхода на поверхность почвы. Затем мицелий проникает в конус нарастания, продвигается вверх, пронизывая узлы и междоузлия, и первое время не вызывает видимого угнетения растений. Ко.времени образования цветочной метелки мицелий гриба достигает ее и обильно " там разрастается, используя богатые питательными веществами ткани, разрушая и вытесняя их. К этому времени мицелий распадается на отдельные клетки, одевающиеся темной оболочкой, — телиоспоры. Вначале телиоспоры двухъядерны, затем ядра сливаются (кариогамия) и образуется диплоидное ядро. Затем происходит редукционное деление ядра и переход гаплоидных ядер в развивающиеся базидиоспоры.
При выколашивании больных растений проса из влагалища верх-
318 Прорастание телиоспор головневых грибов. А- ШШащо; Б Маёо геае; В-Ь. тик; Г-ТШеШШШ
2 _ колос' пшеницы, пораженный головней, 2 — зерна, наполненные спорами ^1^овневые мешочки), 3 - распьшенне спор из Ра^^енного зерна 4 - спора на зерне пшеницы; 5 - прорастание спор на зерне, 6 — внедрение ростков гриба в проростки пшеницы него листа вместо метелки выходит пузыревидное вздутие, состоящее главным образом из головневых спор, покрытое беловатой пленкой, состоящей из остатков мицелия. Пленка легко разрушается и споры высыпаются, особенно во время молотьбы, прилипая к здоровым зернам. Попадая вместе с зернами проса в почву, споры прорастают, и развитие гриба продолжается так, как это указывалось на с. 440. Заражение проростков возбудителями твердой головни пшеницы (рис. 319), стеблевой головни ржи, каменной головни ячменя, головни овса происходит также в почве. У одса. прорастание телиоспоры в базидию и образование диплоидного мицелия наблюдается тотчас после попадания телиоспор под пленки зерен. Мицелий там зимует и после попадания семян в почву заражает проростки. Начальный период развития возбудителя карликовой головни пшеницы — ТШеНа соп1гоьег8а проходит на поверхности почвы. Всходы пшеницы заражаются после выхода их на поверхность почвы.
Увозбудителя пыльной головни; ницы — С/$Щадо 1гШа и ячменя — V. пийа телиоспоры для дальнейшего развитая" должны попасть на рыльца цветков питающих растений, где они прорастают четырехклеточной базидией. Там происходит редукционное деление и образование гаплоидных ядер. Однако базидиоспоры здесь не развиваются, а ди- карион возникает путем перехода ядер из одной клетки базидии в другую либо путем копуляции соседних клеток базидий (рис. 318, В) или клеток соседних базидий (рис. 320, А). Из ставшей двухъядер- ной клетки базидии вырастает дикариотический мицелий, по пыльцевой трубке проникающий в завязь. Однако это не мешает развитию семени, которое внешне кажется нормальным, хотя в тканях его эндосперма и в зародыше находится мицелий паразита. Зараженные семена, попав в почву, нормально прорастают и из них сначала развиваются внешне здоровые растения. Однако по мере дальнейшего развития растения трогается в рост и мицелий гриба, проникающий в росток, распространяющийся в тканях по межклетникам и особенно обильно накапливающийся в конусе нарастания. Затем мицелий проникает в залагающийся Рис. 321. Развитие возбудителя пыльиой головни пшеницы (схема): 2.— рассенвание и перенос спор на цветущие растения, 3 —споры на рыльце цветка, / — ростки грнба проникают в зародыш семени, 5 — зараженное зерно (внешне не отличается от здорового), 6 — срез зараженного зерна под микроскопом (в оболочке виден мицелий грнба —с), 7 —прорастание грибницы колос, обильно там разрастается, разрушает зерна и колоски, в конце концов распадаясь на отдельные телиоспоры. В вышедшем из лиСтового влагалища колосе сохраняются лишь стержень и сильно разрушенные боковые колоски. Остальные, части колоса превращаются в темную пылящую массу телиоспор. Телиоспоры, способные прорастать без периода покоя, переносятся ветром на цветущие в это время колосья, заражая завязи так, как это описано выше (рис. 321). ^".Возбудитель пузырчатой головни кукурузы — II. геае вызывает местные поражения различных органов растений (рис. 322): междоузлий стеблей, листьев, женских и мужских цветков. -Заражаться могут как молодые, так и взрослые растения, однако у взрослых восприимчивы лишь молодые растущие ткани. Телиоспоры возбудителя пузырчатой головни кукурузы — V. геае прорастают четырехклеточной базидией. Базидиоспоры • начинают почковаться еще на базидии, образуя цепочки постепенно отпадающих. клеток (рис. 318, Б). Клетки эти воздушными течениями переносятся на те или иные органы растений кукурузы, где они копулируют, имея разные половые знаки (« + » и «—»). Возникающий после копуляции дикариотичный мицелий и заражает растения. Проникнув в ткани, мицелий разрастается, вызывая местные поражения в виде галлов или вздутий, заполненных телиоспорами паразита. Патологический ■ рост вздутий — опухолей стимулируют образуемые грибом ростовые вещества. Внутри опухоли мицелий обильно разрастается, разрушая ткань опухоли и распадаясь затем на отдельные телиоспоры. В начале образования опухоли масса спор прикрыта эпидермисом, после разрыва которого споры рассеиваются воздушными течениями. Телиоспоры способны тотчас прорастать, вызывая новые заражения растений. Головневые грибы разделяются на два семейства: устиляговые (1ЫПа^тасеае) с разделенной поперечными перегородками четырехклеточной базидией (фрагмобазидия) и тиллециевые (ТШеИасеае) с одноклеточной (не разделенной перегородками) базидией и базидио- спорами, развивающимися на ее конце (рис. 318, Г). В семействе устиляговые наиболее распространен род устиляго — изШадо, у представителей которого довольно крупные тем- ноокрашенные шиповатые, реже гладкие телиоспоры. Грибы, относящиеся к этому роду, поражают главным образом репродуктивные органы. Известно около 350 видов рода С/зШадо, из которых в СССР встречается около 120.
В этом же семействе у грибов рода сфацелотека — ЗрНасеШНеса споровая масса вначале прикрыта оболочкой, состоящей из стерильных гиф и разрушенных тканей субстрата. Позднее оболочка разрывается и находящиеся под ней телиоспоры распыляются. Споры одиночные, коричневв1е или оливковые, чаще шиповатые." В роде около 30 видов. А Б В ~ Г Рис. 323. Споры головневых грибов. А—8ого5роп'ит; Б — ТШеИа\ В — ЦгосузИв; Г—Еп1ку1ота (из С. Р. Шварцман) У рода сороспориум — Зогозропит телиоспоры собраны в клубочки, окруженные студенистой оболочкой из нитевидных гиф, затем исчезающей, после чего клубочки легко распадаются. Телиоспоры разной формы — от шаровидной до угловатой, в массе темно- или светло-красновато-коричневые (рис. 323, А). Известно около 30 видов. В семействе тиллециевые основной род тиллеция — ТШе- Иа, включающий около 80 видов. Телиоспоры довольно крупные, большей частью легко распыляющиеся, округлые или овальные, образуются обычно в завязях, редко в других частях зараженных растений (рис. 323, Б). Споровая масса часто с селедочным запахом. Базидия одноклеточная, на ее вершине сидят четыре серповидные базидиоспоры, часто соединенные копуляционным каналом (рис. 320, Б). Род включает около 80 видов. У рода уроцистис—ИгосувИз телиоспоры чаще формируются в надземных вегетативных органах питающих растений, редко на корнях и в соцветиях. Пораженные части растений искривляются, на них появляются черные полосы и вздутия, сперва прикрытые эпидермисом, который впоследствии разрывается. Телиоспоры соединены в округлые или эллипсоидальные клубочки (спорокучки) по пять — десять клеток, из которых одна-две центральные, более крупные и более темные^ окружены несколькими более мелкими светлыми клетками (рис. 323, В). Прорастают только центральные клетки, остальные прорастать не способны. Базидия короткая, цилиндрическая, на вершине ее развивается пучок из нескольких базидиоспор. В роде около 60 видов. В роде энтилома — ЕШу1ота около 100 видов. Виды этого рода образуют на листьях, стеблях, цветоножках или черешках, редко на корнях растений пятна или вздутия, наполненные телиоспорами. Споры одиночные, от светло-желтых до коричневых, чаще с гладкой, иногда двухслойной оболочкой (рис. 323, Г). О происхождении головневых грибов существуют различные предположения. Ряд учёных считает, что их предков следует искать среди некоторых групп сапротрофных дрожжеподобных организмов из видов леукоспориум (ЬеисозрогШшт), родоспориум (ЮюйозрогШшт) и тил- летария (ТИШапа). Основанием для этого служит сходство цикла развития этих организмов с циклом развития головневых, а также наличие у некоторых видов пряжек, характерных для головневых грибов. Другие ученые считают, что головневые грибы произошли от простейших аурикуляриевых грибов с разделенной перегородками четырехклеточной базидией, развивающейся из пробазидии. Одноклеточная базидия семейства тиллетиевых, возможно, развивалась из многоклеточной базидии, характерной для семейства устиляговые. Между ними есть переходные формы. У некоторых видов устиляговых, например у 11$Ша§о йотезИса, базидиоспоры образуются не по одной из каждой клетки базидии, а почти одновременно в большом числе в виде мутовки базидиоспор. В то же время у ТиЬиг- сша опегМаИв из тиллетиевых базидиоспоры формируются в верхней части базидии, отделенной от остальных частей перегородкой. Как указывалось, у головневых грибов установлена тесная связь •с питающим растением: У большинства видов это проявляется прежде всего в том, что гриб, проникнув в растение, до перехода к спорообразованию в завязях или других частях- мало его угнетает. Пораженные головней растения кустятся несколько сильнее, чем здоровые. У них наблюдается небольшое укорачивание стебля. Действие гриба при поражении большинства хлебных злаков проявляется после проникновения его в репродуктивные органы. При этом колоски почти полностью разрушаются или от них сохраняются лишь осевые, части, а у некоторых видов злаков остается лишь стержень колоса. При сильном заспорении семян хлебных злаков телиоспорами всходы гибнут (например, при поражении пшеницы твердой головней). Поражение пшеницы и других злаков карликовой головней (ТИШТа < соп1гоьег& а) приводит к значительному отставанию больных растений в росте. Вследствие параллельной эволюции головневых грибов и питающих растений у тех и других весьма близки требования к условиям жизни. Например, у возбудителя твердой головни пшеницы температурный оптимум для прорастания телиоспор и заражения растений близок с таковым для прорастания семян пшеницы. Поэтому когда после посева температура почвы близка к благоприятной для растения и паразита (около 12°С), поражение растений бывает особенно сильным. У возбудителей пыльной головни пшеницы и ячменя, заражающих завязи, рассеивание телиоспор приурочено ко времени цветения. Это обеспечивает попадание инфекционного начала на восприимчивые части растений как раз в критический для заражения период. Несмотря на приспособленность к облигатному паразитическому образу жизни, головневые грибы могут расти на синтетических питательных средах, а некоторые из них проходят там полный цикл развития до образования телиоспор. Попавшие в почву телиоспоры головневых грибов сохраняют там жизнеспособность до тех пор, пока не прорастут. Базидии и мицелий довольно быстро разрушаются почвенными микроорганизмами, поэтому значение почвенной инфекции при заражении злаков головней невелико. Главное значение имеет инфекция, находящаяся на семенах или внутри их. Временами головневые грибы и в настоящее время наносят существенный вред хлебным злакам. Наиболее опасны в нащей стране сле- „V • щне головневые грибы. Пшеница. Пыльная головня — 11вШадо ШНа. Твердая или вонючая головня: в северной и средней полосе эту болезнь вызывает ТШеНа сапев, в южной — Т. }оейс1а. Карликовая головня — Т. соп1го- ьегза, местами стеблевая головня — Ь'госузИз 1гШси Овес. Пыльная — 11$Ша§о аюепае и покрытая головня—С/. Рожь. Стеблевая головня—1]госу8И8 оссиЫа. __ .Ячмень. Пыльная головня — 1! зШадо пийа, черная головня — II. тцга, каменная головня — V. Ногйег. Кукуруза. Пузырчатая головня—V. геае, пыльная головня — Зогозрогшт геШапит. Просо. Пыльная головня — ЗрНасёШНеса рапШ-тШасеи Лук. Листовая головня — ИгосувИз сери1ае. Поскольку при поражении головневыми грибами многих злаков уничтожаются все зерна, вред от головневых на этих растениях равен числу пораженных колосьев. Однако иногда вред оказывается несколько выше, так как при сильном заспорении зерна часть всходов погибает, за счет чего уменьшается-число продуктивных растений. Вред от пузырчатой головни кукурузы зависит от числа головневых вздутий на растениях, а также от места их возникновения. Особенно вредоносно появление вздутий на початках и при основании стеблей. Порядок ржавчинные — 11гес1|па1е8 Грибы порядка — паразиты высших растений., широко распространенные в разных широтах. Их насчитывается около 5 тыс. видов. У ржавчинных грибов известны несколько следующих друг за другом спороношений, оканчивающихся образованием покоящихся спор, называемых телейтоспорамиПо своему биологическому значению эти споры соответствуют телиоспорам головневых грибов. Они прорастают в четырехклеточную базидию, клетки которой расположёны одна над другой. Мицелий ржавчинных грибов распространяется по межклетникам тканей зараженных растений, в клетки которых внедряются гаустории. Он содержит капли масла, окрашенные в оранжевый цвет пигментом, близким к каротину. Такие же капли масла имеются в спорах. Пораженные растения покрываются подушечками (пустулами) различных оттенков оранжевого или красно-бурого цвета, поэтому болезнь, вызываемая этими грибами, получила название ржавчины. Цикл развития ржавчинных грибов удобно проследить на примере возбудителя стеблевой, линейной, или черной, ржавчины — РйссШа дгатШз, паразитирующей на барбарисе и на многих хлебныхГ~КОр*~ мовых й дикорастущих злаках. 1 В настоящее время некоторые ученые предложили следующие обозначения спороношений ржавчинных грибов: эции и эциоспоры, урединин и урединиоспоры, телии и телиоспоры. В настоящей книге мы придерживаемся установившейся в нашей стране терминологии. Развитие гриба начинается с прорастания зимующих на соломе телейтоспор и образования базидии ^„базидиоспорами. Перед этим происходит сначала кариогамия (образование диплоидного ядра), а затем редукционное деление. Гаплоидные ядра по одному попадают -^каждую клетку базидии и каждая из них формирует одну базидиоспору, куда й переходят гаплоидные ядра/ Базидиоспоры — маленькие тонкостенные клетки, сидящие на тонких сте- ригмах. При созревании они отбрасываются на некоторое расстояние и затем распространяются воздушными течениями. Для дальнейшего развития базидиоспоры должны попасть на листья барбариса, где формируется весенняя фаза развития возбудителя стеблевой ржавчины.. После проникновения ростка базидиоспоры в ткань листа сначала появляются.оранжевые пятна, ' образованные гипертрофированной тканью листа, пронизанной по межклетникам мицелием гриба с гаусториями, внедрившимися в клетки. Гифы состоят из одноядерных клеток с оранжевыми каплями масла и принадлежат к гаплоидной стадии развития гриба. На хаком мицелии в тканях закладываются два типа спороношений: на верхней поверхности листа пикниды (или спермогонии), на нижней — э ц и- дии (рис. 324). Первоначально оба типа спороношений представляют ~ -собой шаровидный клубок гаплоидных гиф. конечном итоге принимают вид кувшинов, в полости которых от стенок вырастают короткие спсгроносцы, несущие очень мелкие шаровидные споры, называемые пикноспорами или спермациями. Развиваясь, пикниды давят на верхний эпидермис листа, прорывают его.и высовываются наружу в виде коротких гиф, называемых перифизами. К этим гифам приливают пикноспоры (рис. 325). Между ними наружу выступает сахаристая пахучая жидкость с массой " погруженных в нее пикноспор. Эта " " жидкость привлекает насекомых, на брюшке и лапках которых пикноспоры переносятся с одной пикниды на другую. Новых заражений пикноспоры не вызывают, но играют определенную роль в возникновении дикариотического мицелия, т. е. в половом процессе. •
Базидиоспоры ржавчинных грибов раздельнополы. На базидии образуется по две споры разных половых знаков (всего четыре споры). Каждая из них дает гаплоидный мицелий своего, полового знака, кото Возможно также, что на одном и том же листе появляются два спермогония, возникающие в результате заражения базидио- спорами разных половых знаков. Межклеточные мицелии их в толще листа срастаются, образуя дикариотический мицелий, дающий начало двухъядерным гифам, на которых развиваются эцидиоспоры (эци- диальное спороношение). Эцидии первоначально дифференцируются на две части: обращенную к нижнему эпидермису — стерильную, сложенную из крупных, бедных содержимым пузыревидных клеток, и плодущую, сложенную из гиф, богатых цитоплазмой. В цитоплазме появляются двухъядерные клетки, возникающие от слияния двух одноядерных, происходящих в свою очередь от соединения гаплоидных гиф разных половых знаков. Каждая двухъядерная клетка, называемая базальной, начинает отшну-
ровывать на своем конце цепочку эцидио- спор, или весенних спор. При этом оба ядра базальной клетки делятся одновременно и параллельно, и кончик ее отделяется в виде двухъядерной клетки, представляющей материнскую клетку эцидиоспоры. Затем таким же образом отшнуровывается вторая клетка, за ней третья и т. д. Материнская клетка эци- диоспор делится (каждая) на две двухъядерные неравные по объему клетки. У верхней, более крупной, вырабатывается толстая оболочка, и она превращается в эцидиоспору, нижняя — промежуточная клетка — остается тонкостенной и позднее разрушается. • Краевые базальные клетки, располагающиеся кольцом вокруг всего палисадного слоя, также отчленяют цепочки двухъядерных клеток, которые, срастаясь 29 Заказ 552
друг с другом, формируют однослойную оболочку, или перидий. Конечные эцидиоспоры претерпевают такое же изменение, образуя крышу перидия, срастающуюся с его боковыми стенками. Эцидиоспоры, окруженные перидием и его крышей, называются эцидиями. Нарастающие своим основанием базальные клетки в конечном итоге разрывают нижний эпидермис листа и выдвигаются наружу. Крыша перидия разрывается, края его отворачиваются в виде лопастей, и эцидий принимает форму урны, заполненной оранжевыми эцидиоспорами, по мере созревания выпадающими наружу и распространяющимися по воздуху (рис. 326, Л). У некоторых видов ржавчинных грибов перидий не образуется, а „эцидиоспоры, тоже цепочками, сидят на поверхности плоского сплетения. гиф. Такой тип эцидиального спороношения называется цеомой " (рис. 326, 5). Эцидиоспоры округлые, двухъядерные. Заражать барбарис они не могут и для дальнейшего развития должны попасть на листья или стебли злаков, например на пшеницу. Здесь эцидиоспоры прорастают, и ростки их через устьица проникают в ткани, где формируется межклеточный дикариотичный мицелий с гаусториями. На таком двухъядерном мицелии развивается летнее спо- 4)аношение_ гриба — уредоспоры. Каждая уредо- спора сидит на " клетке-ножке (рис. 327). Под давлением уредоспороношений эпидермис листа или стебля разрывается, обнажая желтые, бурые, кирпично-красные или оранжевые подушечки уредоспор — то, что называется ржавчиной. Уредоспоры отрываются от своих ножек, рассеиваются в воздухе и могут снова заражать зла- ^ ки. Прорастая, они развивают диплоидный мицелий, проникающий через устьица в ткани растений. Через несколько дней снова образуются ^уредоспоры, вызывающие новые заражения рас- чтений. Таких новых поколений уредоспор за лето может быть до десяти. Чем быстрее проходит период от заражения до возникновения нового поколения уредоспор, тем сильнее развивается на злаках ржавчина. По мере истощения питательного субстрата (растения) на том же дикариотическом мицелии возникают новые спороношення — телейтоспоры, или зимние споры. У возбудителя стеблевой ржавчины они двухклеточные, сидят на ножке, каждая из клеток первоначально имеет по два ядра, позднее сливающихся в одно диплоидное ядро. Оболочки телейтоспор толстые, темно- бурые, с одной ростковой порой в каждой клетке. При поражении злаков стеблевой ржавчиной к концу вегетации подушечки телейтоспор покрывают стебли" и листовые влагалища черными продольными полосами. Поэтому этот вид ржавчины называется также черной и линейной. Телейтоспоры возбудителя стеблевой ржавчины служат для перезимовки гриба, прорастая лишь после периода покоя. Зимуют они на стерне или соломе (в скирдах), а на диких злаках также на стоящих зимующих стеблях. На соломе, лежащей на земле, телейтоспоры к весне почти всегда погибают от воздействия почвенных микроорганизмов. Весной каждая клетка телейтоспоры прорастает четырехклеточной базидией (рис. 328), куда переходит диплоидное ядро, делящееся затем редукцнонно два раза. Далее образуются базидиоспоры, заражающие барбарис, на котором развиваются пикниды и эцндии (см. с. 448). Таким образом, цикл развития возбудителя стеблевой ржавчины включает следующие типы спороношений: 0 — пикниды с пикноспора- ми, не вызывающими заражения растений, но участвующими в половом процессе (диплоидизации мицелия); I—эцидии с эцидноспорами; II—уредоспороношения (несколько поколений); III — телейтоепоро- ношения; IV — базидии с базидиоспорами (рис. 329).
Гаплоидная фаза начинается с базидий и базидиоспор. В результате заражения растений базидиоспорами развивается гаплоидный ми-
целии и закладываются спороношения, в которвгх пикниды остаются гаплоидными. Диплоидная фаза начинается со слияния двух гаплоидных мице- лиев с разными половыми знаками. На этом мицелии закладываются диплоидные (дикарио- тические) 'эцидиоспоры, заражающие злаки, где развивается несколько поколений уредоспор, а.затем и телейтоспоры. В телейтоспорах происходит кариогамия, а при развитии базидии и переходе туда диплоидного ядра — редукционное де- деление и развитие гаплоидных базидиоспор.
Пикниды и эцидии образуются на барбарисе, уредо- и телейтоспороношения — на злаках. Развитие разных стадий ржавчинных грибов на различных растениях носпт названиегразнохо-
ГО.' 6-Е в Рис. 331. Телейтоспоры ржавчинных грибов семейств'а Ме! ашр5огасеае. А — СгопагНит; Б — МеШтвога; В — Со1еовропит\ Г — СНгувотуха зяинности. Разнохозяинность свойственна " многим ржавчинным грибам, однако среди них есть виды, все стадии развития которых проходят на одном и том же растении. Такие грибы называют однохозяинными. К ним относятся ржавчинные грибы, поражающие подсолнечник, лен, малину и др. У ржавчинных грибов, имеющих все указанные выше стадии, цикл развития называется полным. Однако есть виды, у которых та или иная стадия выпадает. Они относятся к грибам с. неполным циклом развития. Например, у возбудителя ржавчины груши известны только О, I, III и IV стадии, у возбудителя желтой ржавчины злаков неизвестны 0 и I стадии. Порядок ржавчинные грибы — игесНпа1е8 делят на два семейства. Семейство пукциниевые — Риспшасеае. Телейтоспоры одиночные, у немногих — в цепочках, на ножках (рис. 330) с промежуточными клетками. Грибы, относящиеся к этому семейству, паразитируют на покрытосеменных растениях, среди них есть как разнохозяинные, так и однохозяинные виды. Наиболее распространены следующие роды. Род уромицес—1]готусев насчитывает свыше 500 видов. Телейтоспоры одноклеточные (рис. 330, Б), эцидий с перидием. Виды рода часто паразитируют на бобовых растениях (уредо- и телейтостадии) и на молочаях (эцидии). Род пукциния — РисгШа содержит около 2000 видов. Телейто- споры двуклеточные, эцидии с перидием. Паразитируют на злаках, многих видах двудольных. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 296; Нарушение авторского права страницы
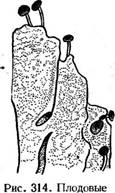 тела РЫеоцепа [а-
фпеа
тела РЫеоцепа [а-
фпеа
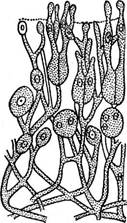
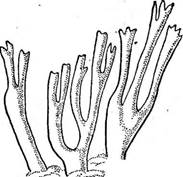

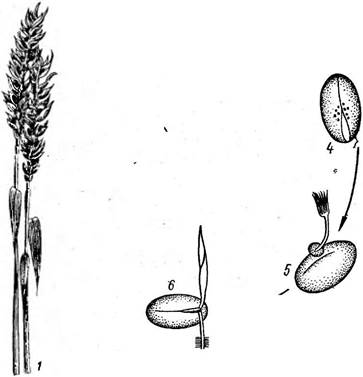 Рис. 319. Развитие твердой головни пшеницы (схема):
Рис. 319. Развитие твердой головни пшеницы (схема):
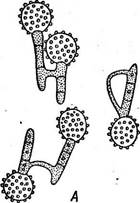
 Рис. 320. Копуляция базидиоспор у головневых грибов. А — род ОзШадо; Б — род ТШеНа
Рис. 320. Копуляция базидиоспор у головневых грибов. А — род ОзШадо; Б — род ТШеНа
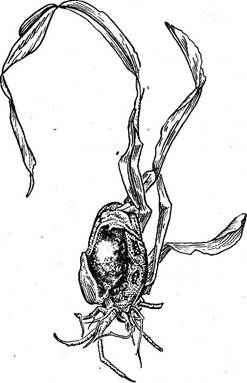 Рис. 322. Пузырчатая головня кукурузы (головневые вздутия на молодых растениях)
Рис. 322. Пузырчатая головня кукурузы (головневые вздутия на молодых растениях)
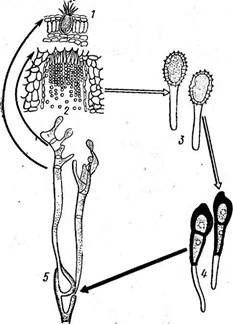 Рис. 324. Цикл развития стеблевой ржавчины злаков— Рисата §гаттв:
/ — пикиида и 2— эцидии на листе бэрбарнса, 3 — уредоспоры и 4 — телейтоспоры на стеблях ■ злаков, 5 — прорастание телейтоспоры в бззи- дию с базидиоспорами
Рис. 324. Цикл развития стеблевой ржавчины злаков— Рисата §гаттв:
/ — пикиида и 2— эцидии на листе бэрбарнса, 3 — уредоспоры и 4 — телейтоспоры на стеблях ■ злаков, 5 — прорастание телейтоспоры в бззи- дию с базидиоспорами
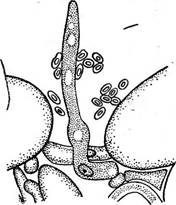
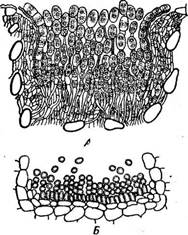 Рис. 326. Типы эцидиев ржавчии- иых грибов. А — эцидий; Б — цеома
Рис. 326. Типы эцидиев ржавчии- иых грибов. А — эцидий; Б — цеома
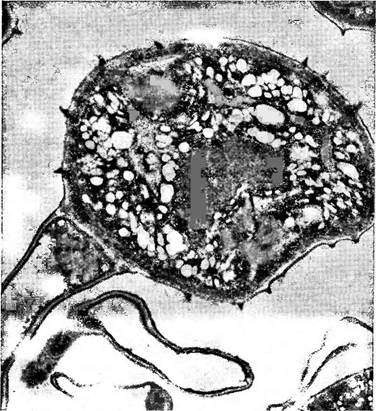 Рис. 327. Уредоспора при увеличении в 3900 раз. В центре видны два ядра (по Ю. М. Плотниковой и М. А. Сассен)
Рис. 327. Уредоспора при увеличении в 3900 раз. В центре видны два ядра (по Ю. М. Плотниковой и М. А. Сассен)
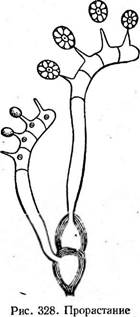 телейтоспор РиссШа фрагмобазидией с базидиоспорами
телейтоспор РиссШа фрагмобазидией с базидиоспорами

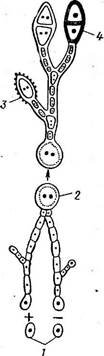 Рис. 329. Схема смеиы ядерных фаз у РиссШа ^гапйшв. Тонким контуром обведена гаплоидная фаза; двойным тонким контуром — дикариофит; жирным контуром— фаза, содержащая диплоидные ядра;
I — базидиоспора. 2 — эци- диоспорг. 3— уредоспора, 4 — телейтоспора
Рис. 329. Схема смеиы ядерных фаз у РиссШа ^гапйшв. Тонким контуром обведена гаплоидная фаза; двойным тонким контуром — дикариофит; жирным контуром— фаза, содержащая диплоидные ядра;
I — базидиоспора. 2 — эци- диоспорг. 3— уредоспора, 4 — телейтоспора

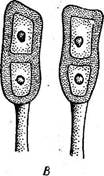 Рис. 330. Телейтоспоры разных родов ржавчинных грибов. А — РНгад- IпШит; Б — Vготусез; В— РиссШа
Рис. 330. Телейтоспоры разных родов ржавчинных грибов. А — РНгад- IпШит; Б — Vготусез; В— РиссШа
