|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Предмет и задачи биохимии(кафедре 110 лет)Стр 1 из 27Следующая ⇒
Билет№1 Билет №2 1)Классификация белков. Структуры белков и методы их определения. По химическому сост.1) Простые белки (протеины) гидролизуются кислотами или щелочами до аминокислот и не дают при гидролизе других органических и неорганических соединений. 1. Альбумины –б плазмы крови с массой40 кДа ( кислые .↑ отрицательный заряд) транспортер для билирубина и жирных кислот. 2.Глобулины – белки, слаб кис. Нейтр. Белок плазмы крови.3. Проламины – белки, растворяются в 60-80%-ном этилового спирта. Пролина. Раст происход 4.Глютелины – белки, которые извлекаются из растительных тканей разбавленными растворами щелочей (0,2%-ный NаОН). 5. Протамины – это белки ↓массы, состоящие на 80% основ св-ва. Аргинин.6. Гистоны –↓белки также основного характера, содержатся в хромосомах клеточных ядер и играют важную роль в образовании структуры хроматина. 2) Сложные белки содерж простетическую группу. 1.Хромопротеины(гемопротеины,магнийпорфирины,флавопротеины) 2.гемопротеины (белок+гем) гем и его произвдные.3 флавопротеины (белок+изоаллоксазиновые производные ФМН и ФАД). Фермент катал ОВР 4.магнийпорфирины 5 нуклеопротеины (белок+нк к-та)6. Липопротеины 7гликопротеины (бел+углевод) 8 металлопротеины (б+ме) фосфопротеины (б+фосфат( 2)Переваривание и всасывание углеводов в ЖКТ Переваривание углеводов С пищей в основном поступают крахмал, гликоген, целлюлоза, сахароза, лактоза, мальтоза, глюкоза и фруктоза, рибоза. Ротовая полость Со слюной сюда поступает кальций-содержащий фермент α-амилаза. Оптимум ее рН 7,1-7,2, активируется ионами Cl–.расщепляет внутренние α1,4-гликозидные связи и не влияет на другие типы связей. крахмал и гликоген = до декстринов и мальтозы. Дисахариды ничем не гидролизуются. Желудок Из-за ↓ рН амилаза инактивируется, некоторое время расщепление углеводов продолжается внутри пищевого комка.Кишечник совместно панкреатическая α-амилаза, разрывающая внутренние α1,4-связи, изомальтаза, разрывающая α1,6-связи изомальтозы, олиго-α1,6-глюкозидаза, действующая на крахмала и гликогена. Пристеночное пищевар:сахаразо-изомальтазный комплекс (рабочее название сахараза) – в тощей кишке гидролизует α1,2-, α1,4-, α1,6-гликозидные связи, расщепляет сахарозу, мальтозу, мальтотриозу, изомальтозу,гликоамилазный комплекс – находится в расщепляет α1,4-гликозидные связи в олигосахаридах, β-гликозидазный комплекс (рабочее название лактаза) – гидролизует β1,4-гликозидные связи между галактозой и глюкозой (лактозу). клетчатка (целлюлоза), которая незначительно расщепляется в толстом кишечнике под действием β -глюкозидазы, выделяемой микробами. Биологическое значение клетчатки очень велико: - создание объема пищи; - усиление перистальтики кишечника и продвижение пищи; - очистка ворсинок тонкого и толстого кишечника; - усиление биосинтеза и секреции ферментов. Всасывание моносахаридов Образовавшиеся в тонком кишечнике моносахариды - глюкоза, фруктоза и галактоза - всасываются по механизму симпорта за счет градиента концентрации катионов натрия, который создает Na+/K+-ATФ – аза. Скорость всасывания отдельных моносахаридов различна: глюкоза -100%, фруктоза - 42, галактоза -110, пентозы - 19. Регуляция синтеза гемма 1. Скорость ↑ гем 2. Основ регул = аминолевулинатсинтаза. гем + белка-репрессора формирует активный репрессорный комплекс, связывается с ДНК и ↓ транскрипцию, мРНК для фермента не образуется и синтез прекращается. количество fe 2+ ↑ синтез аминолевулинатсинтазы. Ионы железа связываются с этим железосвязывающим белком, образуя с ним неактивный комплекс, что ↓синтез фермента. 3. тгипоксия тканей, которая в эритропоэтических тканях ↓синтез фермента. 4. В печени ↑аминолевулинатсинтазы вызывают соединения, ↑ работу микросомальной системы окисления (жирорастворимые токсины, стероиды) – при этом ↑потребление гема для образования цитохрома Р450, ↓внутриклеточную концентрацию свободного гема. => ↓синтеза фермента. Ингибиторы:тяжелые металлы.,глюкоза,гем гемоглобин ↓АЛК синтазы. Стероидные гормоны, нестероидные противовоспалительные средства, антибиотики сульфаниламиды ↑синтазы. Лекарства ↑усвоение гема в системе цитохрома Р450, который важен для продукции данных соединений печенью. Порфирины представляют собой циклические соединения, образованные четырьмя пиррольными кольцами, связанными между собой метенильными мостиками Характерным св-в способность образовывать комплексы с ионами металлов, связывающимися с атомами азота пиррольных колец. Примерами служат железопорфирины, в частности гем, входящий в состав гемоглобина, и магний - содержащий порфирин хлорофилл — пигмент растений, участвующий в фотосинтезе.В природе металлопорфирины связываются с белками = соединения для биологических процессов. A. Гемоглобин — железопорфирины, связанные с белком глобином. Гемоглобины обладают способностью обратимо связывать кислород, они транспортируют этот газ в системе кровообращения Б. Эритрокруорины — железопорфиринопротеины, находящиеся в крови и тканевых жидкостях некоторых беспозвоночных; выполняют такую же функцию, как и гемоглобин. B. Миоглобины—дыхательные пигменты, находящиеся в мышечных клетках позвоночных и беспозвоночных. Билет №3 Гликолиз Гликолиз – это последовательность ферментативных реакций, приводящих к превращению глюкозы в пируват + АТФ.Если О2 мало = ПВК→лакттат. Анаэробный гликолиз – сложный ферментативный процесс распада глюкозы, протекающий в тканях человека и животных без потребления кислорода. Конечным продуктом-молочная кислота. 11 ферментов, протекает в цитоплазме клетки.
2АТФ обарз: 7 и 10 ре-я 2НАДН обрз 6 реак-я (=6АТФ) Затрат НАДН 11 ре-я Всего 8 АТФ Ферменты: гексокиназа+АТФ→глюк-6-Р-изомераза→6-фосфофрукткиназа+АТФ→альдолаза→триозофосфатизомраза→глицеральдегидфосфатдегидрогеназа+НАД+Н2РО4→фосфоглицераткиназа+АДФ+магний→фосфоглицеромутаза→енолаза-Н2О→пируваткиназа+магний→лактатдегидрогеназа+НАДН2 Био значение = образ АТФ. На первых стадиях гликолиза затрачиваются 2 молекулы АТФ (гексокиназная и фосфофрук-токиназная реакции). На последующих образуются 4 молекулы АТФ (фосфоглицераткиназная и пируваткиназная реакции). Таким образом, энергетическая эффективность гликолиза в анаэробных условиях составляет 2 молекулы АТФ на одну молекулу глюкозы., Лимитирующей скорость гликолиза, является фосфофруктокиназная. Вторая реакция, лимитирующая скорость и регулирующая гликолиз – гексокиназная реакция. Кроме того, контроль гликолиза осуществляется также ЛДГ и ее изоферментами. Система фибринолиза. Система фибринолиза 2. Активаторы плазминогена тканевого типа (t-PA, tissue plasminogen activator) и урокиназный (u-PA, урокиназа, urokinase plasminogen activator) – ферменты (сериновые протеазы), превращающие плазминоген в плазмин.тканевой активатор плазминогена (t-PA) выделяется эндотелием, моноцитами, мегакариоцитами,урокиназный активатор плазминогена (u-PA) продуцируется эпителиальными клетками почечных протоков, юкстагломерулярными клетками, фибробластами, макрофагами, эндотелиоцитами. 3. Фактор XII (фактор Хагемана) – контактный фактор, активатор плазминогена и прекалликреина. 4. Прекалликреин – контактный фактор, фактор Флетчера, профермент калликреина, катализирующего образование кининов, но для этого должен сначала активироваться фактором Хагемана (ф.XIIа). 5. Высокомолекулярный кининоген (ВМК, фактор Фитцжеральда) – в кровотоке находится в комплексе с фактором XII, рецептором прекалликреина. Ключевым ферментом= плазмин, гидролизующий фибрин до растворимых продуктов. Активаторы превращения плазминогена в плазмин образуются сосудистой стенкой (внутренняя активация) или тканями (внешняя активация). Билет №4 Билет №5 1)Классификация и биолог роль нуклеиновых кислот Полимеры. НК – высокомолекулярные азотсодержащие органические соединения, структурной единицей которых являются мононуклеотиды, соединенные в полинуклеотидную цепь 3’ – 5’ фосфодиэфирными связями, имеющими сложную структурную организацию и в соединении с белками, определяющими вид, форму, состав, функции живой клетки. Существует два типа НК — дезоксирибонуклеиновые (ДНК) и рибонуклеиновые (РНК). ДНП – преимущественно в ядрах (митохондрии, хлоропласты)РНП - преимущественно вцитоплазме(ядра, ядрышки) Мономерами служат нуклеотиды. Каждый из них содержит азотистое основание, пятиуглеродный сахар (дезоксирибоза — в ДНК, рибоза — в РНК) и остаток фосфорной кислоты. Билет №6 Хромопротеины Для них простетическая часть окрашена.К хромопротеинам относятся гемоглобин, миоглобин, каталаза, пероксидаза, ряд флавинсодержащих ферментов (сукцинатдегидрогеназа, альдегидоксидаза, ксантиноксидаза), цитохромы (гемсодержащие белки) и т.д.Роль– участвуют в физиологических процессах: дыхание клетки, транспорте кислорода и углекислого газа, окислительно-восстановительных процессах. Метаболизм аммиака Источники 1)окислительное дезаминирование аминокислот(основной путь получения) Из организма аммиак выводится почками в виде конечных продуктов азотистого обмена: • мочевины - синтезируется в печени; • аммонийных солей - образуются в почках.
Глутамин - транспортной формой аммиака, нейтральной аминокислотой , легко проник через кл мембраны диффузией .Глутамин поступает в кровь из тканей ,транспортируется в почки и кишечник. В клетках кишечника → 4. В почках глутамин →глутаминазы и расщепляется на глутамат, реабсорбируется и возвращается в клетки тканей, и аммиак Глутаминаза почек активируется при ацидозе; Роль пути выведения: • поддерживает кислотно-щелочной баланс в норме; • защищает организм от потери с мочой ионов Na+ и К+, которые также могут использоваться для выведения избытка анионов.
В мозге восстановительного аминирования α-кетоглутаратапод →глутаматдегидрогеназа→ катализирует реакцию, обратную окислительному дезаминированию глутамата. Из мышц, клеток кишечника избыток азота выводится в кровь в виде аланина .Образование аланина в этих органах где подвергается непрямому дезаминированию. Выделившийся аммиак обезвреживается в процессе синтеза мочевины, а пируват включается в глюконеогенез или ОПК. Глюкоза из печени поступает в ткани и в процессе гликолиза окисляется до ПВК. Образование аланина в мышцах, его перенос в печень и перенос глюкозы в обратном направлении составляют глюкозоаланиновый цикл. 7. В печени аммиак обезвреживается путем связывания с СО2 и образования карбамоилфосфата.Реакцию ката карбамоилфосфатсинтетаза I, использует 2 моль АТФ. Фермент локализован в митохондриях гепатоцитов. Продукт реакции -карбамоилфосфат - включается затем в орнитиновый цикл Кребса-Гензелейта для синтеза мочевины. Регуляция Поддержание осмотического давления. В почках альдостерон увеличивает реабсорбцию Nа+ в канальцах и, вследствие осмотических явлений, реабсорбцию воды. Происходит облегчение транспорта К+ и Н+ в противоположном направлении антидиуретический гормон (АДГ, вазопрессин) – гормон нейрогипофиза. АДГ усиливает реабсорбцию воды в дистальных отделах почечных канальцев, что приводит к торможению диуреза. Естественным стимулом секреции АДГ является возбуждение осморецепторов.
Билет №7 1)Гемоглобин Гемоглобин входит в состав группы белков гемопротеины, подвидом хромопротеинов Небелковой частью их является гем – структура, включающая в себя порфириновое кольцо (состоящее из 4 пиррольных колец) и иона Fe2+. Железо + порфириновым кольцом двумя координационными и двумя ковалентными связями.Координ.число=2 Пат типы. HbS – гемоглобин серповидно-клеточной анемии.(остальные буквы, кроме а,р,f Пат.формы. HbCN(Fe3+)цианметHb (fe2+) HbS(Fe2+)сульфоHb Билет №8 Гликогенез Гликоген-главная форма запасания глюкозы (печень, мышцы) Гликогенез- бх реакция, синтеза гликогена из глюкозы Синтез: глюкоза→(ф.гексокиназа)-затрата АТФ →глюкозо6фосфат→(ф.фосфоглюкомутаза. обратим) глюкозо-1-фосфат (ф. глюкозо1фосфатуридилтрансфераза) +уридинтрифосфат(УТФ)→УТФ-глюкоза + пирофосфат Удф глюкоза+гликоген (n остаток (больше 4))→(ф. гликогенсинтаза) гликоген (n+1 остатков) Амило-α-1,4-α-1,6-гликозилтрансфераза ("гликогенветвящий"фермент) переносит фрагмент (6 остатков глюкозы) на соседнюю цепь, образуя α-1,6-гликозидную связь (рис. 11). Гликогенсинтаз а образует α-1,4-гликозидные связи и удлиняет гликогеновую цепочку, присоединяя первый углеродный атом УДФ-глюкозы к четвертому атому углерода концевого остатка гликогена. Глюкозо-1-фосфат -уридилтрансфераза-ключевой фермент. Регуляция: Инсулин- ↑ синтеза гликогена – активация гликогенсинтазы и стимуляция ее синтеза, что облегчает превращение излишков глюкозы в гликоген, Глюкагон ↑ глюкозы крови:↑ мобилизацию гликогена через активацию гликогенфосфорилазы, стимулируя глюконеогенез – ↑ работы ферментов пируваткарбоксилазы, фосфоенолпируват-карбоксикиназы, фруктозо-1,6-дифосфатазы. Адреналин стимул гликогенфосфорилазы, Глюкокортикоиды ↑глюкозу крови = ↓ перехода глюкозы в клетку,↑ глюконеогенез – ↑ синтез ферментов пируваткарбоксилазы, фосфоенолпируват-карбоксикиназы, фруктозо-1,6-дифосфатазы. 3)Глюкокортикоиды Билет №9 Метаболизм ацетил-КоА Из:б окисления жирных кислот, расщепление аминокислот,окисление глюкозы до ПВК,оксилительное декарбоксилирование. При алкогольном опьянении.Одновременно из-за относительной недостаточности оксалоацетата избыток "алкогольного" ацетил-SКоА не успевает окислиться в цикле трикарбоновых кислот и перенаправляется на синтез кетоновых тел.. Учавсвует: окисление в ЦТК, синтез кетоновых тел,синтез холестерина,биосинтез жирных кислот. Ацетил-КоА может окисляться в цикле трикарбоновых кислот.= 12 молекул АТФ на каждую молекулу ацетилкоэнзима А. Ацетил-КоА для синтеза жирных кислот, при этом требуется достаточное количество энергии (АТФ). При недостатке энергии, при нарушении обмена углеводов и липидов синтез холестерина через мевалоновую кислоту нарушается и происходит образование кетоновых тел с их накоплением в тканях и выделением с мочой (кетозы). Мевалоновоя кислота является предшественником холестерина. При нормальной энергетической обеспеченности ацетоацетил-КоА полностью превращается в печени в мевалоновую кислоту, затем в холестерин. При ↓ углеводов, сахарном диабете, нарушении обмена липидов происходит↑кетоновых тел, развиваются кетозы. Кетоновые тела: активированная мевалоновая кислота распадается на ацетоуксусную кислоту и ацетил-КоА: Ацетоуксусная кислота частично восстанавливается до (β-оксимасляной кислоты, а частично декарбоксилируется, образуя ацетон: II. Если CH3-CO-KoA образуется много и недостаточно энергии для синтеза жира= активированная ацетоуксусная кислота→ свободная ацетоуксусная кислота:→в β-оксимасляную кислоту или ацетон. Кетоновые тела синтезируются в печени и поступают в кровь. В нормальных условиях β-оксимасляная кислота, ацетоуксусная кислота окисляются в тканях мышц и являются источником энергии. Окисление ненасыщенных жирных кислот Синтез высших жирных кислот происходит из ацетил-KoA в цитозоле. Из митохондрий через чулночный механизм в цитозоль. Фермент ацетил-КоА-карбоксилаза, содержащий биотин, катализирует АТФ-зависимый синтез малонил-КоА из ацетил-КоА и СО2: Синтез холестерина. Белки соединительной ткани. В состав соединительной ткани входит вода (63%.) и плотные вещества (37%). Главным компонентом плотного вещества является белок коллаген (85%), а также эластин (4,4%) и ретикулин (0,5%). Альбумины и глобулины составляют 0,6%. Другими компонентами плотного вещества являются гликозоаминогликаны (ГАГ -3,5) и липиды (2,8%). Экстрактивные вещества составляют 3,2%, а неорганические - 0,5% от общего количества. Коллаген. Характеристика первичной структуры тропоколлагена, т.е. аминокислотного состава. глицина приходится 33%, аланина 11%, пролина и оксипролина 21%, лизина и оксилизина 1-3%. первичной структуры =↓ метионина, нет цистеин и триптофан. Вторичную и третичную структуру тропоколлагена = трехцепочная молекула. б)Эластин. В состав первичной структуры проэластина входят необычные аминокислоты (десмозин, лизиннорлейцин). в)Свойства белков соединительной ткани. Существенными свойствами являются эластичность, прочность, волокнистость, плохая растворимость в воде, устойчивость к денатурации, плохая перевариваемость в желудочно-кишечном тракте,↓ антигенность, а также биологическая неполноценность этих белков из-за особого аминокислотного состава. Синтез и распад коллагена. Синтез тропоколлагена (проколлагена) осуществляется из протоколлагена с участием лизина и пролингидрокилаз, ↑р02, ↑ Fе+2, витамина С и альфа-кетоглутарата в среде.Гидроксилирование лизина и пролина→гликозилирование→«сборка» 3-х цепочной молекулы тропоколлагена→ суперспирализация , с участием ГАГ , в фибриллы коллагена. Распад коллагена осуществляется специфическим ферментом коллагеназой, → «разрушает» 3 пептидные цепи на ¼ расстояния от С-конца между глицином и лейцином → гидролизуются пептидами. 4 типа коллагена: 1 тип - коллаген сухожилий, связок, II тип -хрящей, III тип - кровеносных сосудов, кожи, кишечника и IV тип - коллаген базальных мембран. Протеогликаны. Белки промежуточного вещества. В их состав входят гликозоаминогликаны (ГАГ-95%) и белки - альбумины и глобулины (5%) ГАГ . Структура ГАГ (димеры, повторяющиеся). Различают ГАГ: гиалуроновая кислота, хондроитинсерная кислота, хондронтин-4-сульфат, хондронтин-6-сульфат, дерматансульфат, кератансульфат, гепарансульфат, гепарин. + уроновые кислоты (глюкуроновая или идуроновая) Биологическая роль ГАГ: активное связывание воды, двухвалентных катионов (Са++ и Мg++), участвуют в образовании фибрилл и деградации хиломикронов (гликопротеидов), обладают антикоагулянтным свойством (гепарин), факторы агрегации клеток. Синтез и распад ГАГ осуществляется в ЭР фибробластов с участием УТФ, ацетил КоА, ФАФС и других метаболитов. Распад ГАГ регулируется ферментами лизосом фибробластов. В регуляции синтеза коллагена и ГАГ (протеогликанов) =гормоны и биологически активные вещества. Ингибиторами синтеза =глюкокортикоиды надпочечников. Активаторов =минералокортикоиды, андрогены (тестостерон) и соматомедин. Билет №10 1)Уровни организации структуры ферментов. Мультиферментные комплексы. Ферменты – это глобулярные белки, состоящие из одной или нескольких полипептидных цепей, организованных в 3-х мерную структуру. По химическому строению различают простые и сложные (имеющие небелковую часть - кофактор). Кофакторы – это в-ва, способствующие действию ферментов, или необходимые для его осуществления. Роль· изменение 3-ой структуры белка и создание комплементарности между ферментом и субстратом, стабилизация молекулы фермента в пространстве; · непосредственное участие в реакции в качестве еще одного субстрата. Кофакторы =коферменты и простетические группы. Добавочную группу, прочно связанную с белковой части= простетической группой; легко отделяющуюся от апофермента и способную к самостоятельному существованию= коферментом. В составе фермента выделяют области, выполняющие различную функцию: 1. Активный центр – комбинация аминокислотных остатков (обычно 12-16), обеспечивающая непосредственное связывание с молекулой субстрата и осуществляющая катализ. Аминокислотные радикалы в активном центре могут находиться в любом сочетании, при этом рядом располагаются аминокислоты, значительно удаленные друг от друга в линейной цепи. В активном центре выделяют два участка: якорный (контактный, связывающий) – отвечает за связывание и ориентацию субстрата в активном центре, каталитический – непосредственно отвечает за осуществление реакции. У ферментов, имеющих в своем составе несколько мономеров, может быть несколько активных центров по числу субъединиц. Также две и более субъединицы могут формировать один активный центр. У сложных ферментов в активном центре обязательно расположены функциональные группы кофактора. 2. Аллостерический центр– центр регуляции активности фермента, который пространственно отделен от активного центра и имеется не у всех ферментов. Связывание с аллостерическим центром какой-либо молекулы (называемой активатором или ингибитором, а также эффектором, модулятором, регулятором) вызывает изменение конфигурации белка-фермента и, как следствие, скорости ферментативной реакции. Аллостерические ферменты являются полимерными белками, активный и регуляторный центры находятся в разных субъединицах. В качестве такого регулятора может выступать продукт данной или одной из последующих реакций, субстрат реакции или иное вещество Мультиферментные комплексы В мультиферментном комплексе несколько ферментов прочно связаны между собой в единый комплекс и осуществляют ряд последовательных реакций, в которых продукт реакции непосредственно передается на следующий фермент и является только его субстратом. Благодаря таким комплексам значительно ускоряется скорость превращения молекул. Например:пируватдегидрогеназный комплекс (пируватдегидрогеназа), превращающий пируват в ацетил-SКоА,α-кетоглутаратдегидрогеназный комплекс (в цикле трикарбоновых кислот) превращающий α-кетоглутарат в сукцинил-SКоА, комплекс под названием "синтаза жирных кислот" (или пальмитатсинтаза), синтезирующий пальмитиновую кислоту. Углеводный обмен наработку НАДФН, необходимого для микросомального окисления и синтеза жирных кислот и холестерола из глюкозы. Липидный обмен Связь углеводного и липидного обмена происходит на уровне дигидроксиацетонфосфата (ДАФ) и ацетил-КоА. ДАФ, при гликолизе →восстанавливается в глицеролфосфат→ синтез триглицеридов. Ацетил-КоА образуется при окислительном декарбоксилирования пирувата →образования жирных кислот, кетоновых тел, холестерина, желчных кислот, стероидных гормонов. В ходе большинства из этих синтетических процессов используется НАДФН(Н+), поставщиком =окислительная ветвь пентозофосфатного пути превращения углеводов. Белковый обмен ПФП поставляет также рибозо-5-фосфат, используемый для биосинтеза нулеиновых кислот. Таким образом на уровне данного метаболита прослеживается взаимосвязьуглеводного и нуклеинового обмена. Метаболитов( пируват и оксалоацетат), образ в процессе обмена углеводов →в реакциях трансаминирования → в аминокислоты α-аланин и аспартат→в синтез белка. ПВК →щавелевоуксусную и a-кетоглютаровую (кислоты, из → реакции аминирования и переаминирования → аспарагиновая и глютаминовая аминокислоты. Возможен и обратный процесс превращения аминокислот в глюкозу. Глюкогенные= глицин, α-аланин, серин, цистеин, треонин, метионин, валин, аспарагиновая кислота, аспарагин, глутаминовая кислота, глутамин, аргинин, гистидин, пролин. Безазотистые остатки →в– пируват, кетоглутарат, сукцинил-КоА, фумарат, оксалоацетат→через пируват и оксалоацетат →в глюконеогенез. Но, как и при превращениии липидов в углеводы, эти процессы также носят ограниченный характер.. 3)Биохимический состав мочи. Диагностическое значение определения нормальных и патологических компонентов. 1. Мочевина - 30г. главный органический азотсодержащий компонент мочи. Экскреция мочевины зависит от состояния желудочно-кишечного тракта, функций печени, почек, интенсивности метаболизма. ↑мочевины - гиперуриурия ↑употреблением белка с пищей, носит алиментарный характер, гиперуриурия при всех заболеваниях, сопровождающихся ↑распадом белков тканей (лихорадочные состояния, кахексия, гипертиреоз, сахарный диабет и др.), при приеме некоторых лекарственных средств (гормонов). ↓мочевины, выделяемой с мочой, гипоуриурия, =заболеваний и токсических поражений печени, сопровождающихся печеночной недостаточностью, заболеваний почек с нарушением их фильтрационной способности, применении инсулина. 2. Мочевая кислота – 0.7 г. конечный продукт катаболизма пуриновых нуклеотидов. ↑мочевой кислоты (гиперурикурия), при лейкемии, полицитемии, гепатитах, подагре, а также при лечении аспирином и кортикостероидами.Гиперурикурия = гиперпродукцией в организме мочевой кислоты → ↑распада тканевых структур или генетических нарушений отдельных ферментов - синдром Леша-Нейхана и др. → незначительной растворимости в воде мочевая кислота= камни в нижних отделах мочевых путей. 3. Креатинин - 18-32г конечным продуктом азотистого обмена. Образуется в мышечной ткани из креатина. Креатинин →мочу через клубочковой фильтрации → активной канальцевой секреции. Гиперкреатининурия =приеме ↑мясной пищи, некрозе мягких тканей (мышц), синдроме длительного раздавливания, тяжелой мышечной работе, после снятия кровоостанавливающего жгута, лихорадочных состояниях, пневмонии. Гипокреатининурия =хроническом нефрите с уремией (почечная недостаточность), мышечной атрофии, дегенерации почек, лейкемии, в старческом возрасте.. 4. Общий азот 1.1г-глицерин, гистидин,глутамин, аланин, серин. Гиперазотурия = ↑распаде тканевых белков (сахарный диабет, тиреотоксикоз и др.), гипоазотурия - при ↓белка в питании, при нарушении выделительной функции почек. Гиппуровая кислота-нарушение функций печени. Безаазотистые органические компоненты – щавеливая,молочная, и имонная,маслян,валериан,янтар. Билет №11 1)Современные представления о механизме ферментативного катализа. ЭНЕРГЕТИЧЕСКИЙ БАРЬЕР РЕАКЦИИ – кол-во энергии, которое необходимо молекуле, чтобы вступить в химическую реакцию. ЭНЕРГИЯ АКТИВАЦИИ - кол-во энергии, которое необходимо сообщить молекуле для преодоления ЭБР. 1. В спонтанной реакции – 18 ккал/моль 2. При использовании химического катализатора – 12 ккал/моль 3. В присутствии фермента – 5 ккал/моль ) Гидролиз белков в желудке – 20 ккал/ моль, а в присутствии пепсина – 12 ккал/моль роль ферментов =↓ энергии активации. Ферменты не изменяются во время химической реакции катализируют как прямую, так и обратную реакцию Действуют в ничтожно малых концентрациях Активность зависит от температуры Не влияют на величину К равновесия Не изменяют свободную энергию (ΔG) Молекулярный механизм действия ферментов Сближение и ориентация– активный центр фермента связывается с субстратом Напряжение и деформация- «эффект дыбы», растягивание субстрата, индукция соответствия S и Е. кислотно-основный катализ– в активном центре фермента есть группы специфичных аминокислотных остатков= донорами или акцепторами протонов. мощные катализаторы многих органических реакций. Ковалентный катализ –ф + субстратами= помощи ковалентных связей очень нестабильные фермент-субстратные комплексы→продукты реакции. Тип "пинг-понг " – фермент + субстратом А, отбирая у него химические группы →продукт→ ф+субстрат В, + эти химические группы. Примером = переноса аминогрупп от аминокислот на кетокислоты - трансаминирование. Тип последовательных реакций – к ф+ последовательно субстраты А и В= "тройной комплекс→ катализ. Продукты последовательно отщепляются от фермента. 3. Тип случайных взаимодействий – субстраты А и В + к ф в любом порядке, неупорядоченно→катализа →отщепляются. В основу современной теории положена теория Михаэлеса и Ментена. 1 этап: фермент-субстратного комплекса.Диффузия с+ф =принципом комплиментарности, + с активным центром =ф-субстратный комплекс. Реагенты связаны слабыми связями, Эта стадия непродолжительна, зав концентрации субстрата и от скорости диффузии его к активному центру. ется эффект концентрирования субстрата на поверхности фермента – эффект ориентации. 2 этап: преобразование первичного ФСК в 1 или несколько активированных. длительность зависит от величины энергии активации данной реакции. разрыв старых связей и образование новых, энергия активации значительно ↓ По продолжительности =лимитирующей для всего процесса.эффект «дыбы 1) 3 этап: отделение продуктов от активного центра фермента и диффузия их в окружающую среду. Эта стадия непродолжительна, ее скорость определяется скоростью диффузии продуктов в окружающую среду. 2) КАТАЛ – такое количество фермента, которое может осуществить превращение 1 моль субстрата за 1 сек. Катал = Моль/с, мМоль/с, мкМоль/с, нМоль/с Билет №12 1) Кинетика ферментативного катализа 1. Стереоспецифичность – катализ только одного из стереоизомеров, например:специфичность к L- или D-аминокислотам специфичность к цис- и транс-изомерам. Например, аспартаза реагирует только с транс-изомером – фумаровой кислотой, но не с малеатом (цис-изомер). 2. Абсолютная специфичность – фермент производит катализ только одного вещества. Например, расщепление мочевины уреазой. 3. Групповая специфичность – катализ субстратов с общими структурными особенностями, т.е. при наличии определенной связи или химической группы: например, наличие пептидной связинапример, наличие ОНгруппы: алкогольдегидрогеназа окисляет до альдегидов одноатомные спирты (этанол, метанол, пропанол). 4. Относительная групповая специфичность – превращение субстратов с некоторыми общими признаками. Все сводится к комплементарному взаимодействию фермента и субстрата. Наличие субстратной специфичности объясняют две гипотезы: 1. Теория Фишера (модель "жесткой матрицы", "ключ-замок") – активный центр фермента строго соответствует конфигурации субстрата и не изменяется при его присоединении. Эта модель хорошо объясняет абсолютную специфичность, но не групповую. 2. Теория Кошланда (модель "индуцированного соответствия", "рука-перчатка") – подразумевает гибкость активного центра. Присоединение субстрата к якорному участку фермента вызывает изменение конфигурации каталитического центра таким образом, чтобы его форма соответствовала форме субстрата. Зависимость от температуры: ↑ температура, тем ↑скорость, но оптимум 40 градусов (после денатурация белка) Зависимость от ph: от 6-до 8 . специфична. Заивисиость от концентрации субстрата.Теоретический график зависимости скорости ферментативной реакции от концентрации субстрата при постоянной концентрации фермента. ( Михаэлиса – Ментен) 1. Концентрация субстрата равна величине констансты Михаэлиса ([S] = Km). скорость реакции V = ½ Vmax ↑величины Кm означает ↓ сродства фермента к субстрату. 2. Концентрация субстрата значительно больше Km ([S] >> Kmчто скорость реакции максимальна 3. Концентрация субстрата значительно меньше Km ([S] << Km). В этом случае, знаменатель уравнения мало изменяется при изменении [S], а величина скорости реакции V прямо пропорциональна [S] (график линеен). 2)Гликогенолиз. Регуляция концентрации глюкозы крови. Гликоген печени расщепляется ↓концентрации глюкозы в крови, прежде всего между приемами пищи. Через 12-18 часов голодания его запасы в органе полностью истощаются. В мышцах количество гликогена ↓физ нагруз. Из-за отсутствия в них глюкозо-6-фосфатазы находящийся в клетках отрицательно заряженный эфир моносахарида не способен преодолеть цитолемму и выйти в кровь, что позволяет использовать гликоген только для собственных нужд.В гликогенолизе непосредственно участвуют три фермента:Фосфорилаза гликогена разрывает α-1,4-гликозидные связи с отщеплением глюкозо-1-фосфата. Фермент работает до тех пор, пока до точки ветвления (α-1,6-связи) не останется 4 остатка глюкозы. · α(1,4)-α(1,6)-Глюкантрансфераза – энзим, переносящий фрагмент трисахарида на другую цепь с образованием новой α-1,4-гликозидной связи. При этом на прежнем месте остается один остаток глюкозы и "открытая" действию катализатора доступная α-1,6-гликозидная связь.Амило-α-1,6-глюкозидаза, ("деветвящий" фермент) гидролизует последнюю с отрывом свободной (нефосфорилированной) глюкозы. В результате возникает цепь без ветвлений, вновь служащая субстратом для фосфорилазы. Аденилатциклазанеактив→(адреналин,глюкогон)актив→+атф=цАМФ→цамфзависимая протеинкиназа(н)→(а)→киназа фосфорила б(н)→+атф и магний(а)→фосфорилаза б (н)+4 атф-4адф=(а)→гликоген n+н3ро4→глюкозо-1-фосфат→глюкозо6фосфат→глюкоза. факторами гормональной регуляции =яглюкагон, адреналин, глюкокортикоиды, соматотропный гормон с одной стороны, и инсулинс другой ↑синтеза гликогена – активация гликогенсинтазы и стимуляция ее синтеза, что облегчает превращение излишков глюкозы в гликоген, активация пентозофосфатного пути – индукция синтезаглюкозо-6-фосфат- дегидрогеназы и 6-фосфоглюконатдегидрогеназы, усиление липогенеза – вовлечение глюкозы в синтез триацилглицеролов Глюкагон ↑глюкозы крови: ↑ мобилизацию гликогена через активацию гликогенфосфорилазы, стимулируя глюконеогенез – повышение работы ферментов пируваткарбоксилазы,фосфоенолпируват-карбоксикиназы,фруктозо-1,6-дифосфатазы. Адреналин -активируя мобилизацию гликогена – стимуляция гликогенфосфорилазы, Глюкокортикоиды ↑ глюкозу крови за счет подавления перехода глюкозы в клетку, стимулируя глюконеогенез – ↑ синтез ферментов пируваткарбоксилазы,фосфоенолпируват-карбоксикиназы,фруктозы-1,6-дифосфатазы 3.Иммуноглобулины сыворотки крови. Иммуноглобулины G – основные иммуноглобулины сыворотки, осуществляют гуморальную защиту организма от бактерий и вирусов и их растворимых токсинов (антигенов). Активно транспортируются через плаценту. У детей должная концентрация достигается только к 1,5-2 годам. Иммуноглобулины А осуществляют местный иммунитет на слизистых поверхностях дыхательных путей (носовой и бронхиальный секрет) и кишечного тракта. Они присутствуют в женском молозиве, слезе, слюне. После рождения количество секреторного IgA (на слизистых) достигает уровня взрослых уже на 2-3 неделе жизни, сывороточного IgA – только к 14-15 годам. Иммуноглобулины М появляются в процессе формирования иммунного ответа, являясь первичными антителами. Вскоре после рождения их уровень нарастает, достигает максимума к 9 месяцу жизни, после чего снижается и восстанавливается только к 20-30 годам. Иммуноглобулины Е вырабатываются плазматическими клетками и участвуют в аллергических реакциях организма. Иммуноглобулины D не имеют четкой сформированной функции. Предполагается, что они регулируют активность других иммуноглобулинов.
Билет №13 1. Регуляция активности ферментов. 1. Влияние закона действия масс (обратимая р.трансаминирования: аланин + ὰ-кетоглутарат↔пируват +глутамат). 2. Изменение кол.фермента (феномен индуцированного синтеза фермента; конститутивные ферменты: зависит от наличия продукта реакции, катализируемой данным ферментом, продукт реакции вызывает торможение синтеза фермента в результате репрессии). 3. Проферменты (зимогены в активную форму). 4. Химическая модификация фермента(обратимая ковалентная и нековалентная). 5.Аллостерическая регуляция (ингибирование по принципу обратной связи; сопровождается изменением сродства фермента к субстрату без изменения максимальной скорости реакции). 6. Явление компартментализаци (разделить несовместимые метаболические процессы; разъединяет ферменты с субстарами). Метаболизм этанола. Основным местом метаболической трансформации этанола – печень. Этанол дегидрируется алкогольдегидрогеназой в этаналь (ацетальдегид)+надн2 в в цитоплазме→митохондрии альдегиддегидрогеназой переводится в ацетат+надн2. Уксусная кислота в реакции, катализируемой ацетат-КоА-лигазой (тиокиназой) в присутствии АТФ, превращается в ацетил-КоА (ацетил-СоА).→синтез жирных кислот→синтез триглицеридов. Наряду с цитоплазматической алкогольдегидрогеназой в метаболизме этанола принимают ограниченное участие каталаза и "индуцибельная" микросомальная алкогольоксидаза. Скорость трансформации этанола в печени лимитируется главным образом активностью алкогольдегидрогеназы. Другим лимитирующим фактором является наличие НАД+. Максимальная скорость реакции наблюдается даже при небольших концентрациях этанола→уровень этанола в организме ↓с постоянной скоростью (расщепление этанола — реакция нулевого порядка). «Энергетическая ценность» этанола составляет 29,4 кДж/г (7 ккал/г). →алкогольные напитки обеспечивают организм значительной частью энергоресурсов (особенно при алкоголизме). Билет №14 Система антикоагулянтов. Система представлена клетками ретикулоэндотелиальной системы, гепатоцитами и гуморальными факторами. РЭС и гепатоциты удаляют из кровотока активированные факторы свёртывания, включая фибриноген. Система антик. тормозит ктивацию процесса свёртывания крови и разностороннее влияние на фибринолиз. Естественные (эндогенные) антикоагулянты разделяют на первичные и вторичные. Первичные образуются в тканях и в форменных элементах крови. Они всегда присутствуют в плазме Вторичные - образуются в процессе свётрывания крови и фибринолиза в результате протеолитического действия ферментов на свои субстраты. 1)Ингибитор пути тканевого фактора (tissue factor pathway inhibitor, TFPI) синтезируется эндотелиальными клетками, тромбоцитами, моноцитами, фибробластами, гладкомышечными клетками и кардиомиоцитами. функция – ингибирование фактора VIIа, Xa. Кофактором TFPI является протеин S. 2)Выходя из зоны тромбин взаимодействует с тромбомодулином,теряет прокоагулянтнуюактивность, т.е. способность образовывать фибрин и активировать XIII, и приобретает антикоагулянтные свойства (тромбиновый парадокс). Комплекс тромбин-тромбомодулинактивирует антикоагулянтный протеин С, который подавляет работу PAI-1 (ингибитор активатора плазминогена), и который также, при помощи протеина S, ингибирует факторы Vа и VIIIа. Протеины С и S синтезируются в печени при участии витамина К. Функции протеина S: кофактор TFPI (ингибитор пути тканевого фактора), подавляющего TF-VIIa и ф.Xa. Функции активного протеина C (APC) заключаются в том, что он: инактивирует PAI-1, является ингибитором факторов Vа и VIIIа, которые стали доступными для ингибирования после эффекта TFPI (устранилось связывание их с Xa и XIa). 3)Антитромбин III синтезируется в печени, кофактор 4)гепарин. Функцией- инактивация тромбина (ф.IIa) и фактора Xa, на факторы IXa, XIa, XIIа, калликреин, комплекс ТФ-VIIа.
Билет №15 Билет №16 1)Гидролиз нуклеиновых кислот в ЖКТ. Клиническое значение определения мочевой кислоты в крови и моче. Распад нуклеиновых кислот происходит в тонком кишечнике гидролитическим путем →панкреатических нуклеаз. Рибонуклеаза гидролизует только РНК, освобождая мононуклеотиды и олигонуклеотиды. Дезоксиры-бонуклеаза →Mg2 + или Mn2 + и специфически гидролизует ДНК, →к динуклеотид, олигонуклеотидов и мононуклеотидив. Полный гидролиз →действием диэстераз, →слизистой оболочке кишечника. Освобожденные мононуклеотиды расщепляются под → неспецифических фосфатаз или нуклеотидаза до нуклеозидов и фосфорной кислоты. Различают эндонуклеазы, разрывают внутренние межнуклеотидные связи в молекуле ДНК и РНК→деполимеризации нуклеиновых кислот с образованием олигонуклеотидов, и экзонуклеазы, катализирующих гидролитическое отщепления конечных мононуклеотидов от ДНК или РНК. Мононуклеотиды распадаются до конечных продуктов обмена гидролитическим или фосфоролитичним путем (в первом случае для разрыва связей используется вода, во втором - фосфорная кислота). При расщеплении аденозина под действием аденозиндезаминазы образуется инозин→ нуклеозидфосфорилазы-гипоксантин. Ксантиноксидаза - флавопротеинов, содержащий молибден и железо, окисляет гипоксантин в ксантин → мочевую кислоту. окислителя =молекулярный кислород восстанавливается до H2O2, а каталаза разлагает →H2O и O2. Гуанозин под действием нуклеозидфосфорилазы →в гуанин, →гуаниндезаминазы - на ксантин→окисляется в мочевую кислоту. В организме человека мочевая кислота является конечным продуктом расщепления пуринов, она выделяется с мочой. ↑ ее в крови - гиперурикемия-вызывает отложение урата натрия в виде кристаллов в тканях, особенно в суставах и хрящах→подагрой= артритов. Отложения мочевой кислоты →камни в почках, →к повреждению почек. Пиримидиновые азотистые → разрушением пиримидинового кольца. Конечными продуктами распада пиримидиновых азотистых → CO2, NH3, ß-аланин и ß-аминоизомасляна кислота. ß-Аланин → синтеза ансерину и карнозина, а →коэнзима А. ß-Аминоизомасляна → метил-малонат: судьба КоА- ↑оказывается также у больных раком. ↑мочевой кислоты :Лишний вес. Неумеренность в еде.Малое потребление жидкости. Злоупотребление алкоголем.Заболевания ↑ м.к-ты: Болезни суставов.↑ давление, ССС,Сахарный диабет. ↓ мочевой кислоты могут стать следствием низкопуриновой диеты. →Патология Коновалова — Вильсона.Патология Фанкони.Патология Ходжкина. Патологии канальцев почек.Мочевая кислота ↓в крови при беременности. обширными ожогами, опухолевыми заболеваниями, врожденными патологиями печени и при приеме отдельных медикаментов. Билет №17 Билет №18 1. Пути превращения глюкозо-6-фосфата в клетке. 1)Пентозофосфатный путь (прямое окисление УВ) (в цитозоле кл) 2 фазы: • окислительная фаза (окисление глюкозо-6-фосфата и последующее окислительное декарбоксилирование продукта) необратимо окисляется в пентозу - рибулозо-5-фосфат, и образуется восстановленный НАДФН. • неокислительная фаза (превращение пентозофосфатов с образованием исходного глюкозо-6-фосфата) рибулозо-5-фосфат обратимо превращается в рибозо-5-фосфат и метаболиты гликолиза. Пентозофосфатный путь обеспечивает клетки рибозой для синтеза пуриновых и пиримидиновых нуклеотидов и гидрированным коферментом НАДФН, который используется в восстановительных процессах. 2) Гликолиз. Глюкозо-6-фосфат превращается в фруктозо-6-фосфат (фермент – глюкозо-6-6фосфат изомераза) и далее до образования малата или в ЦТК Билет №19 Метаболическая функция 1.Почки осуществляют ряд биохимических синтезов: • синтез эритропоэтина, стимулирующего продукцию эритроцитов, • 2.синтезируется активатор плазминогена урокиназа. • гидроксилирование 25-оксикальциферола и превращение его в 1,25-_диоксикальциферол, регулирующий кальциевый обмен, 3.в почках происходит синтез глюкозы (глюконеогенез) из органических кислот (лактата, пирувата). Основное значение глюконеогенез в почках имеет при полном голодании – при этом образуется до 50% всей глюкозы, находяящейся в крови. Билет №20 Билет№21 Билет№ 22 1)ОКисление жирных кислот в клетке. Расчет количества АТФ 1.Распад жира на глицерин и жк 2.ЖК транспортируется альбумином по крови в клетку Ацильные остатки →через внутреннюю мембрану митохондрий →специального переносчика, карнитин → остаток высшей жирной кислоты переносится с коэнзима А на карнитин → ацилкарнитина: →карнитин-ацилкарнитин-транслоказной системы →через мембрану внутрь митохондрии и в матриксе →карнитин-ацилтрансферазы ( Е2) ацильный остаток →внутримитохондриальный коэнзим А. → ацил-КоА; →ступенчатому циклическому окислению → одного цикла b-окисления радикал жирной кислоты укорачивается на 2 атома углерода, а отщепившийся фрагмент выделяется в виде ацетил-КоА.
β-окислении = 8 молекул ацетил- S КоАПоследний → ЦТК, при его окислении в одном обороте →3 НАДН, 1 ФАДН2 и 1 ГТФ, 7=12 молекулам АТФ. 8 молекул ацетил-S-КоА →8×12=96 молекул АТФ. Гормоны гипофиза. Его называют “дирижером гормонального оркестра организма”, т.е. он оказывает влияние на все остальные железы. Гипофиз состоит из 3-х долей: передняя, средняя, задняя. Уменьшают: соматостатин, кортизол, тироксин и трийодтиронин (по механизму обратной отрицательной связи). Механизм действия.Аденилатциклазный. Мишени и эффекты↑синтез белков, фосфолипидов и нуклеиновых кислот,↑васкуляризацию щитовидной железы,↑ рост и пролиферацию тиреоидных клеток, ↑захват йода и его включение в тиреоглобулин, активирует все стадии образования трийодтиронина и тироксина. АКТГ (адренокортикотропный гормон). По химической природе – полипептид. Необходим для нормального функционирования надпочечников, в особенности корковой части. Гормоны коры надпочечников образуются из холестерина. АКТГ способствует накоплению холестерина в коре надпочечников, а также глюкозы. В процессе окисления глюкозы (пентозо-фосфатный цикл) образуется НАДФН2 , который необходим для синтеза кортикостероидов. При избытке АКТГ развивается болезнь Иценко-Кушинга (гипергликемия, гиперпигментация, гипертония, гипернатриемия). 3) Диагностическое значение исследования ферментов: ЛДГ, КК, АСТ, АЛТ АСТ(аспарат-аминотрансфераза) ↑ при инфаркте миокарда, тромбоз легочной артерии, гепатиты(вирусный,алкогольный,токсический), цирроз, рак печени, забрюшинная флегмна,мышченая дистрофия. АСТ ↓ при некротических процессов в печени, после поворного гемодиализа,дифецит вит В6. Если при разрыве печени АлАт и АсАт ↓, а билирубин ↑илинорм, → неблагоприятным прогностическим признаком. Лабораторным тест - исследование активности изоферментов в сыворотке крови, в частности изоферментов ЛДГ. В сердечной мышце ЛДГ1 и ЛДГ2, а в ткани печени – ЛДГ4 и ЛДГ5. острым инфарктом миокарда в сыворотке крови ↑ЛДГ1 и отчасти ЛДГ2. Изоферментный спектр ЛДГ в сыворотке крови при инфаркте миокарда напоминает изоферментный спектр сердечной мышцы. паренхиматозном гепатите в сыворотке крови ↑ДГ4 и ЛДГ5 и ↓ЛДГ1 и ЛДГ2. БИЛЕТ №23 1)Метаболизм кетоновых тел. Кетоновые тела, являясь продуктами жирового обмена, выполняют в организме функцию энергетического субстрата, т.е. источника энергии. К кетоновым телам относят b-гидроксимасляную кислоту, ацетоуксусную кислоту и, как конечный (тупиковый) продукт – ацетон (диметилкетон). Синтезируются кетоновые тела в печени (в гепатоцитах) из ацетил-КоА. При выраженном углеводном голодании (белковый и белково-жировой рацион с недостатком углеводов), при нарушениях обмена углеводов (сахарный диабет) в результате клеточного голодания ↑распад и использование жиров, → накоплению ацетил-КоА. →в синтез кетоновых тел. ↑кетоновых тел в крови →развитию кетозов и кетоацидозов. Для синтеза кетоновых тел используется 3 ацетил-КоА. При конденсации двух молекул ац.-КоА образуется ацетоацетил-КоА; затем добавление третьей молекулы приводит к образованию b-гидрокси-b-метил-глютарил-КоА. При этих реакциях освобождается две молекулы HS-КоА. Полученная молекула b-гидрокси-b-метил-глютарил-КоА под действием лиазы распадается на ацетоуксусную кислоту и ацетил-КоА. Все указанные реакции протекают в матриксе митохондрий гепатоцитов. В цитоплазме клеток ацетоуксусная кислота при участии НАД-зависимой дегидрогеназы (НАДФ*Н2) может восстанавливаться до b-гидроксимасляной кислоты или спонтанно декарбоксилироваться с образованием ацетона (диметилкетона).
· В норме кетоновые тела идут на : цикл кребса, синтез холестерина,синтез жирных кислот,ФЛ,синтез аминокислот. · 2) Гормоны щитовидной железы. Синтезирует гормоны двух типов: 1. Иодсодержащие тироксин и трииодтиронин (производные тирозина) 3)Белки соединительной ткани. Коллаген. Характеристика первичной структуры тропоколлагена, т.е. аминокислотного состава. глицина приходится 33%, аланина 11%, пролина и оксипролина 21%, лизина и оксилизина 1-3%. первичной структуры =↓ метионина, нет цистеин и триптофан. Вторичную и третичную структуру тропоколлагена = трехцепочная молекула. б)Эластин. В состав первичной структуры проэластина входят необычные аминокислоты (десмозин, лизиннорлейцин). в)Свойства белков соединительной ткани. Существенными свойствами являются эластичность, прочность, волокнистость, плохая растворимость в воде, устойчивость к денатурации, плохая перевариваемость в желудочно-кишечном тракте,↓ антигенность, а также биологическая неполноценность этих белков из-за особого аминокислотного состава. Синтез и распад коллагена. Синтез тропоколлагена (проколлагена) осуществляется из протоколлагена с участием лизина и пролингидрокилаз, ↑р02, ↑ Fе+2, витамина С и альфа-кетоглутарата в среде.Гидроксилирование лизина и пролина→гликозилирование→«сборка» 3-х цепочной молекулы тропоколлагена→ суперспирализация , с участием ГАГ , в фибриллы коллагена. Распад коллагена осуществляется специфическим ферментом коллагеназой, → «разрушает» 3 пептидные цепи на ¼ расстояния от С-конца между глицином и лейцином → гидролизуются пептидами. 4 типа коллагена: 1 тип - коллаген сухожилий, связок, II тип -хрящей, III тип - кровеносных сосудов, кожи, кишечника и IV тип - коллаген базальных мембран. Протеогликаны. Белки промежуточного вещества. В их состав входят гликозоаминогликаны (ГАГ-95%) и белки - альбумины и глобулины (5%) ГАГ . Структура ГАГ (димеры, повторяющиеся). Различают ГАГ: гиалуроновая кислота, хондроитинсерная кислота, хондронтин-4-сульфат, хондронтин-6-сульфат, дерматансульфат, кератансульфат, гепарансульфат, гепарин. + уроновые кислоты (глюкуроновая или идуроновая) Биологическая роль ГАГ: активное связывание воды, двухвалентных катионов (Са++ и Мg++), участвуют в образовании фибрилл и деградации хиломикронов (гликопротеидов), обладают антикоагулянтным свойством (гепарин), факторы агрегации клеток. Синтез и распад ГАГ осуществляется в ЭР фибробластов с участием УТФ, ацетил КоА, ФАФС и других метаболитов. Распад ГАГ регулируется ферментами лизосом фибробластов. В регуляции синтеза коллагена и ГАГ (протеогликанов) =гормоны и биологически активные вещества. Ингибиторами синтеза =глюкокортикоиды надпочечников. Активаторов =минералокортикоиды, андрогены (тестостерон) и соматомедин. БИЛЕТ №24 1) Синтез триглицеридов и фосфолипидов . Триглицериды синтезируются в стенке кишечника, в печени и жировой ткани (в адипоцитах). Синтез триглицеридов в стенке кишечника может происходить из моноглицерида (из 2-моноацилглицерола) и 2активных жирных кислот (остатки жирных кислот в комплексе с ацилпереносящим энзимом – S-КоА), или из глицерина и 3 молекул активных жирных кислот + АТФ.Глицерин фосфорилируется с + АТФ → глицерофосфата (фермент глицеролкиназа), →глицеролфосфатацилтрансферазы взаимодействует с 2 ацилкоэнзима-А (например, с пальмитил-КоА→фосфатидная кислота (3-фосфо-1,2-диацилглицерол) + ацилкоэнзимом-А → триглицерид, свободный HS-КоА и остаток ортофосфорной кислоты. Печень – основной орган, где идет синтез жирных кислот из продуктов гликолиза. Основной путь синтеза триглицелидов в печени из жирных кислот и глицерофосфата. Глицерофосфат→печень поступает из гидролиза жиров и при восстановлении диоксиацетонфосфата (из гликолиза) при помощи восстановленного НАДФ (НАДФ Н+Н) . Фосфолипиды синтезируются во всех клетках, содержащих ядро. Биологическое окисление – это совокупность ОВ превращений различных веществ в живых организмах.ОВР, протекающие с изменением степени окисления атомов вследствие перераспределения электронов между ними. Типы процессов биологического окисления: 1) аэробное (митохондриальное) окисление предназначено для извлечения энергии питательных веществ с участием кислорода и накоплении её в виде АТФ. Аэробное окисление называется также тканевым дыханием, поскольку при его протекании ткани активно потребляют кислород. 2) анаэробное окисление – это вспомогательный способ извлечения энергии веществ без участия кислорода. Анаэробное окисление имеет большое значение при недостатке кислорода, а также при выполнении интенсивной мышечной работы. 3) микросомальное окисление предназначено для обезвреживания лекарств и ядов, а также для синтеза различных веществ: адреналина, норадреналина, меланина в коже, коллагена, жирных кислот, желчных кислот, стероидных гормонов. 4) свободнорадикальное окисление необходимо для регуляции обновления и проницаемости клеточных мембран. Основным путём биологического окисления является митохондриальное, связанное с обеспечением организма энергией в доступной для использования форме. Источниками энергии для человека являются разнообразные органические соединения: углеводы, жиры, белки. В результате окисления питательные вещества →до СО2 и Н2О (при распаде белков также образуется NH3). Выделяемая при этом энергия– АТФ. Микросомальное и свободнорадикальное окисление. не связано с синтезом АТФ. Механизм данного типа окисления субстрата кислородом предусматривает такое взаимодействие субстрата (S) с молекулярным кислородом, при котором один атом кислорода включается в окисляемый субстрат, другой – в молекулу воды. → возникает гидроксильная группа →гидроксилированием.SН + О2 + А∙Н2 → S-ОН + Н2О + Аде SН – окисляемый субстрат; А∙Н2 – донор водорода (аскорбиновая кислота или НАДФ∙Н2).Ферменты, =гидроксилазами, либо оксигеназами. =активном центре ионы металлов с переменной валентностью (Fe, Cu). При растирании ткани фрагменты цитоплазматической сети самопроизвольно замыкаются в пузырьковидные структуры, называемые микросомами, Группа ферментов микросом =циклическую цепь транспорта электронов и протонов, источником →НАДФ∙Н2. Цитохром Р450 с катионом железа (Fe3+ ↔ Fe2+) + субстратом и кислородом → активация и окисление + НАДФ∙Н2. цитохром Р450 отдаёт молекуле кислорода четыре электрона→один из атомов кислорода внедряется по связи С-Н молекулы окисляемого субстрата, а другой – восстанавливается с образованием воды.Биологическая роль микросомального окисления:1. Синтез различных веществ. синтез адреналина и норадреналина в хромаффинной ткани; пигмента меланина из тирозина в коже, радужке и сетчатке глаза; основного белка соединительной ткани – коллагена. Ферменты микросом участвуют в образовании ненасыщенных жирных кислот; желчных кислот и стероидных гормонов надпочечников из холестерина, лейкотриенов из арахидоновой кислоты.2. Обезвреживание различных токсических веществ в печени. ксенобиотиками. → токсические вещества становятся водорастворимыми, →не накапливаются в клетке, → выводятся с мочой. Р450 – это способность его белка изменять свою конформацию в ответ на появление в организме того или иного ксенобиотика, обеспечивая тем самым эффективное взаимодействие с ним. За счёт такой приспособляемости цитохром Р450 является универсальным ферментом детоксикации, способным взаимодействовать почти с любыми соединениями. Единственной требование к окисляемому субстрату – он должен быть неполярным, поскольку цитохром Р450 находится в липидном слое мембран. 3) Калликреин-кинопоказ и ренин-ангиотензиновая системы. 1. Принимает участие во внутреннем механизме образования протромбиназы, благодаря активации фактора XIa; 2. Обеспечивает через фактор XIIa и калликреин взаимосвязь между внешним и внутренним путем образования протромбиназы, активируя факторыVII и XI. 3. Активирует фибринолиз. 4. Участвует в регуляции основных биологических функций сосудистой стенки. В частности, брадикинин расширяет просвет периферических и коронарных артерий↓ артериальное давление ↑проницаемость капилляров. 5. Ферменты кининовой системы активируют С1-компонент комплемента и проренин, а также стимулируют активацию нейтрофилов либо непосредственно, либо через высвобождение брадикинина. 6. Кинины принимают участие в возникновении воспаления, обеспечивая местное развитие его основных признаков; 7. Под влиянием кининов происходит усиление секреции IL-1, TNFa, IL-8, а также простагландинов и лейкотриенов. 8. Кинины оказывают влияние на продукцию и секрецию оксида азота (NO), расширяющего кровеносные сосуды и являющегося мощным дезагрегантом тромбоцитов, активатором фибринолиза и медиатором в ЦНС. 9. Под воздействием брадикинина ↑сокращение гладкой мускулатуры бронхов и других органов и стимулируется болевой эффект. Брадикинин обладает инсулиноподобным действием, способствует усвоению глюкозы периферическими органами, модулирует передачу нервных импульсов в ЦНС. РААС-Секреция ренина в ЮГК регулируется четырьмя основными влияниями. 1. величиной давления крови в приносящей артериоле, ↓ растяжения активирует, а ↑ — подавляет секрецию ренина. 2. регуляция секреции концентрации натрия. ↑натрия в моче дистального канальца, ↑уровень секреции ренина. 3. симпатическими нервами, ветви которых заканчиваются на ЮГК, медиатор норадреналин через бета-адре-норецепторы стимулирует секрецию ренина. 4 отрицательной обратной связи, включаемой уровнем в крови других компонентов системы — ангиотен-зина и альдостерона, а также их эффектами — содержанием в крови натрия, калия, артериальным давлением, концентрацией простагландинов в почке, образующихся под влиянием ангиотензина.Кроме почек образование ренина происходит в эндотелии кровеносных сосудов многих тканей, миокарде, головном мозге, слюнных железах, клубочковой зоне коры надпочечников. ренин →расщепление альфа-глобулина плазмы крови — ангиотензиногена, образующегося в печени. →в крови образуется малоактивный декапептид ангиотензин-I, который в сосудах почек, легких и других тканей подвергается действию превращающего фермента (карбоксикатепсин, кининаза-2), отщепляющего от ангиотензина-1 две аминокислоты. →октапептид ангиотензин-II обладает =стимуляцией клубочковой зоны коры надпочечников, секретирующей альдостерон, Ангиотензин-II, • вызывает сужение артериальных сосудов,• активирует симпатическую нервную систему как на уровне центров, так и способствуя синтезу и освобождению норадреналина в синапсах, ↑ сократимость миокарда,↑ реабсорбцию натрия и ↓ клубочковую фильтрацию в почках,• способствует формированию чувства жажды и питьевого поведения. Таким образом, ренин-ангиотензин-альдостероновая система участвует в регуляции системного и почечного кровообращения, объема циркулирующей крови, водно-солевого обмена и поведения.
БИЛЕТ №25 БИЛЕТ №26 БИЛЕТ №27 Тромбина фактора Виллебранда – рецепторы GPIb,фибриногена – рецепторы GPIIb/IIIa,неактивных и активных форм факторов свертывания XI, X, IX, VIII, V, тромбоксана,коллагена – рецепторы GPIa/IIa,АДФ – рецепторы P2Y2 и P2Y12,серотонина, гистамина. Также в тромбоцитах имеется сократительный белок тромбостенин (ретрактозим) - белок подобный актомиозину, примыкает к внутренней стороне мембраны и связан с внутриклеточным доменом рецептора GPIIb/IIIa. Сосудистая стенка секреция в кровь факторов свертывания, антикоагуляции и фибринолиза, наличие на мембране эндотелия рецепторов для факторов гемостаза,связывание тромбоцитов с коллагеном базальной мембраны. Неповрежденный эндотелий АДФаза,(стимулятор их агрегации),простациклины (PgI), Тканевой активатор плазминогена, Ингибитор фактора (ф.VIla-TF) и, как следствие активацию фактора X. На мембране рецепторы для тромбина и протеина С , участвующие в антикоагулянтной активности. Поврежденный 1. Тканевой фактор ( фактор III) 2. Ингибитор активатора плазминогена 3. Фактор Виллебранда – очень крупный выполняющий функции прикрепления тромбоцита к субэндотелиальному коллагену в поврежденной стенке сосуда (адгезия) через тромбоцитарный рецептор GPIb, связывания и защиты фактора VIII в кровотоке от активированного протеина С, доставка фактора VIII в зону повреждения (т.к. сам фактор Виллебранда здесь задерживается). Билет №28 1)Дезаминирование аминокислот. Виды дезаминирования. Прямое и непрямое дезаминирование. Дезаминирование АК — реакция отщепления α-аминогруппы от АК,А) Прямое дезаминирование Прямому дезаминированию повергаются глу, гис, сер, тре, цис. Существует 4 видова прямого дезаминирования АК: окислительное;внутримолекулярное;восстановительное; гидролитическое. Окислительное дезаминирование: 1. Аэробное прямое окислительное дезаминирование катализируется оксидазами D-аминокислот (D-оксидазы) в качестве кофермента использующими ФАД, и оксидазами L-аминокислот (L-оксидазы) с коферментом ФМН. В организме человека эти ферменты присутствуют, но практически неактивны. 2. Анаэробное прямое окислительное дезаминирование существует только для глутаминовой кислоты, катализируется только глутаматдегидрогеназой, превращающей глутамат в α-кетоглутарат. Фермент глутаматдегидрогеназа имеется в митохондриях всех клеток организма (кроме мышечных). Внутримолекулярное дезаминирование:для гистидина. Реакцию катализирует гистидаза (гистидин-аммиаклиаза). Эта реакция происходит только в печени и коже Б)Непрямое дезаминирование аминокислот идет в 2 этапа: 1 этап(трансаминирование) заключается в обратимом переносе NH2-группы с аминокислоты на кетокислоту с →новой аминокислоты и новой кетокислоты при участии ферментов аминотрансфераз. В качестве кетокислоты-акцептора ("кетокислота 2") в организме обычно используется α-кетоглутаровая кислота, которая → глутамат ("аминокислота 2").→трансаминирования свободные аминокислоты теряют α-NH2-группы и →кетокислоты→ кетоскелет катаболизирует специфическими путями →цикл трикарбоновых кислот и тканевое дыхание→СО2 и Н2О. При необходимости (например, голодание) углеродный скелет глюкогенных аминокислот может →синтеза глюкозы в глюконеогенезе. В этом случае количество аминотрансфераз ↑под влиянием глюкокортикоидов. Второй этап – дезаминирование. →глутаминовая кислота . → глутаматдегидрогеназой.→ в митохондриях печени, аммиак →синтеза мочевины→ удаляется с мочой. В эпителии канальцев почек реакция необходима для удаления аммиака в процессе аммониегенеза. Так как НАДН используется в дыхательной цепи и α-кетоглутарат вовлекается в реакции ЦТК, то реакция активируется при дефиците энергии и ингибируется избытком АТФ и НАДН. Обе стадии непрямого дезаминирования обратимы, что обеспечивает как катаболизм аминокислот так и возможность образования практически любой аминокислоты из соответствующей α-кетокислоты 2) Классификация витаминов, общие свойства и групповая характеристика. Витамин В12 , участие в обмене веществ. ВИТАМИНЫ -низкомолекулярные органические вещества, которые являются регуляторами биохимических процессов, не синтезируются в организме, поэтому должны поступать с пищей. Свойства-они не синтезируются в живых организмах -не накапливаются -не являются источниками энергии;-не включаются в структуру тканей. Классификация:Витаминоподобные (холин,инозит, липоевая кислота, убихинон,оротовая, линолевая,арахидоновая кислоты) § Жирорастворимые Вит.А (антиксерофтальмический) ретинол Вит.D (атирахитический) кальциферол Вит.Е (антистерильный) токоферол Вит.К (антигеморрагический) нафтохинон § Водорастворимые В1 (антиневритный) тиамин В2 (витамин роста) рибофлавин В3 (антидерматитный) пантотеновая кислота В6 (антидерматитный) пиридоксин В12 (антианемический) цианкобаламин РР (антипеллагрический) никотинамид Вс (антианемический) фолиевая кислота Н (антисеборейный) биотин С (антискорбутный) аскорбиновая кислота Р (капилляроукрепляющий) биофловоноиды Групповая характеристика: повышение реактивности организма=регуляция цнс,обмена веществ, трофика, обмен са и р= B 1,2,PP,ACDE 2)антигеморраргические= регул гемостаза=С P K 3)антианемические=кроветвор=CEB12BC 4)антиинфекционные=выработка антителзащитных свойсв=САЕ 5)регул зрен=острот зрен=АСВ2 Витамин В12 (кофак-метилкобаломин) - транспорт метильных групп; - участвует в транспорте водорода; - превращение рибозы в дезоксирибозу; - участвует в синтезе метионина. {Т.е., кобаламин нужен для обмена белков и нуклеиновых кислот.} Недостаток кобаламина: анемия, поражение нервной системы, снижение кислотности желудочного сока. Суточная потребность в кобаламине 0,003 мг. Источники: печень, рыба, яйца, молоко, + синтезируется микрофлорой. (в дрожжах – нет) Недостаток кобаламина может возникать при заболеваниях желудка, в котором вырабатывается внутренний фактор Касла (транс Билет №29 Билет №30 1)Биосинтез мочевины Диагностическое значение определения в крови и моче. Весь цикл мочевинообразования можно представить следующим образом: На первом этапе синтезируется карбамоилфосфат в результате конденсации ионов аммония, двуокиси углерода и фосфата (поступающего из АТФ) под действием фермента карбамоилсинтетазы. Карбамоилфосфат - это метаболически активная форма аммиака, используемая в качестве исходного продукта для синтеза ряда других азотистых соединений. На втором этапе мочевинообразования происходит конденсация карбамоилфосфата и орнитина с образованием цитруллина; реакцию катализирует орнитинкарбамоилтрансфераза. →цитруллин превращается в аргинин в результате двух последовательно протекающих реакций. Первая из них, энергозависимая, сводится к конденсации цитруллина и аспарагиновой кислоты с →аргининосукцината (эту реакцию катализирует аргининосукцинатсинтетаза). Аргининосукцинат →аргинин и фумарат при участии другого фермента - аргининосукцинатлиазы. На последнем этапе аргинин расщепляется на мочевину и орнитин под действием аргиназы. СО2 + NH3 + Аспартат + 3 АТФ + 2 Н2О → Мочевина + Фумарат + 2 (АДФ + Н3Р04) + АМФ + H4P2O7 В реакциях орнитинового цикла расходуются четыре макроэргических связи 3АТФ на каждый оборот цикла. Однако процесс превращения аминокислот в безазотистые остатки и мочевину имеет пути компенсации энергозатрат: при включении фумарата в ЦТК на стадии дегидрирования малата образуется NADH, → синтез 3 молекул АТФ при окислительном дезаминировании глу-тамата в разных органах также образуется NADH, соответственно - ещё 3 молекулы АТФ. ↑ мочевины в крови употреблении в пищу чрезмерного количества белка диете, бедной ионами хлора обезвоживании организма:. приеме некоторых лекарственных препаратов: сульфаниламидов, левомицетина, тетрациклина, гентамицина, фуросемида чрезмерном катаболизме белка: лейкозе, паренхиматозной желтухе, тяжелых нарушении выведения мочевины, связанного с заболеваниями почек и мочевыводящих путей: хронические заболевания почек (гломерулонефрит, пиелонефрит); обструкции мочевыводящих путей (опухоли мочевыводящих путей, предстательной железы, почечно-каменная болезнь); острая и хроническая почечная недостаточность; нарушении выведения мочевины при заболеваниях и состояниях, не связанных с заболеваниями почек и мочевыводящих путей: недостаточность деятельности сердца, острый инфаркт миокарда; сахарный диабет с кетоацидозом, аддисонова болезнь и другие заболевания. ↓ содержания мочевины в крови наблюдается редко: при беременности (вследствие физиологической гидремии); при диете с низким содержанием белка и высоким содержанием углеводов, голодании; при парентеральном введение жидкостей (вследствие гипергидратации);после гемодиализа; ↓катаболизме белков;при нарушении всасывания в кишечнике, целиакии;↑утилизации белка (в поздние сроки беременности, у детей до 1 года, при акромегалии); при врожденной недостаточности или отсутствии ферментов, участвующих в орнитиновом цикле мочевинообразования;при особенно тяжелых поражениях печени (печеночная недостаточность), вызванных, в частности, отравлением фосфором, мышьяком и другими гепатотропными ядами; остром некрозе печени, печеночной коме, декомпенсированном циррозе, гепатитах (известно, что печень обладает большими функциональными резервами, способность ее к дезаминированию и синтезу мочевины сохраняется при исключении из процессов обмена до 85% ее ткани). ↑мочевины с мочой наблюдается при: злокачественной анемии (вследствие отрицательного азотистого баланса);лихорадке;после приема некоторых лекарственных препаратов (салицилатов, хинина, передозировке тироксина и др.);гиперпротеиновой диете; гиперфункции щитовидной железы; ↓мочевины с мочой наблюдается: при диете с низким содержанием белка и высоким содержанием углеводов;при приеме СТГ, тестостерона, инсулина, анаболических гормонов (положительный азотистый баланс);в период выздоровления; при заболеваниях почек и почечной недостаточности любого происхождения;при паренхиматозной желтухе, острой дистрофии печени, прогрессирующем циррозе печени (вследствие нарушения образования мочевины); при врожденной недостаточности или отсутствии ферментов, участвующих в синтезе мочевины; при токсемии. 2) Эйкозаноиды. Тромбоксан -Вызывает агрегацию тромбоцитов Оказывает сосудосуживающее действие Простациклин -Активирует синтез эндотелия сосудов Вызывает дезагрегацию тромбоцитовАктивирует фибринолиз ЛейкотриеныАктивирует аллергические реакции ет имунные реакции анафилактические реакции Сокращение гладких мышц дыхательных путей,пищеварительного тракта,сосудосуживающие действие .Эйкозаноиды действуют как локальные биорегуляторы путём связывания с мембранными рецепторами в непосредственной близости от места их синтеза как на синтезирующие их клетки (аутокринное действие), так и на соседние клетки (паракринное действие). В некоторых случаях их действие опосредовано цАМФ и цГМФ. Билет№31 1)Синтез креатина. Диагностическое значение определения в крови и моче. В синтезе принимают участие три аминокислоты: аргинин, глицин и метионин. 1 стадия – биосинтез гуанидинацетата – осуществляется в почках при участии глицин-амидинотранс-феразы (аргинин+глицин) 2стадия синтеза креатина протекает в печени при участии гуанидинацетатметилтрансферазы Креатин подвергается фосфорилированию с образованием креатин-фосфата, который после дефосфорилирования (необратимая реакция) превращается в креатинин, выделяющийся с мочой. Устойчивое повышение уровня креатинина вкрови, как и возрастание концентрации мочевины в ней, указывает на нарушение функции почечного фильтра. ↓ уровня креатинина в плазме (сыворотке) кровикоррелирует с условленным возрастом уменьшением мышечной массы, оно наблюдается при беременности, наибольшей выраженности в первом и втором триместре. зависит от характера питания. ↑ выведения креатинина с мочой отмечается при большой физической нагрузке (мышечной работе), лихорадочных состояниях, острых инфекционных заболеваниях (прикоторых происходит усиление цитолиза, т.е. повышение распада белков клеток), тяжело протекающем воспалении легких, деструктивном поражении других паренхиматозных органов, в особенностисопровождающемся выраженной недостаточностью функции печени. Гиперкреатинемия наблюдаетсяпри некрозе или атрофии скелетных мышц (травме мышц, мышечной дистрофии, дерматомиозите, полиомиелите, боковом амиотрофическом склерозе, врожденной амиотонии, миастении, голодании), непроходимости кишечника, ожогах,переломах, инфекциях, эндокринных нарушениях (сахарном диабете, гипертиреозе, синдроме Иценко-Кушинга, акромегалии, евнухоидизме), декомпенсации функции сердечно-сосудистой системы, почек, ревматоидном артрите, лейкозах Билет №32 Билет №33 1)Распад гемоглобина. Основные продукты распада, место их образования и пути выведения. Происходит в мононуклеарных фагоцитах (в селезенке и печени). Гемоглобин+гаптоглобин→(гемоксигеназа)+надфр2+2о2-н2о-надф=вердоглобин→+железо+витамин С(фердоглобинредуктаза)биливердин(зеленый пигмент)→(билевердинредуктаза)+2надфн-надф(кл Купфера)билирубин непрямой(коричнево-красный пигмент) связан с альбумином (или свободный)→2УДФПК-2УДФ(глюкоронид-трансфераза)(гематоцит)билирубин прямой +2 глюк-кислоты→желчь(-2 глюк кислоты)→кишечник,билирубин+4H→восстановленный→мезобилирубин→+4Н мезобилиноген(уробилиноген)бесцветный→1) 10%мезобилин→почки→моча→уробилин 2)мезобиин→печень→обезвреживание→желчные дипирролы→либо в кал либо→кровь→почки→моча→моно и дипирролы 3)мезобилин→-4Н→стеркобилиноген (толстая кишка) →либо кал(стеркобилин) либо→вены→кровь→почки→стеркобилиноген мочи→стеркобилин. В кишечнике билирубин подвергается восстановлению под действием микрофлоры до мезобилирубина и мезобилиногена (уробилиногена). Часть последних всасывается и с током крови вновь попадает в печень, где окисляется до ди- и трипирролов. При этом при здоровой печени в общий круг кровообращения и в мочу мезобилирубин и уробилиноген не попадают, а полностью задерживаются гепатоцитами. Оставшаяся в кишечнике часть пигментов ферментами бактериальной флоры толстого кишечника восстанавливается до стеркобилиногена и выделяется из организма, окрашивая кал. Незначительное количество стеркобилиногена через геморроидальные вены попадает в большой круг кровообращения, отсюда в почки и выделяется с мочой. На воздухе стеркобилиноген и уробилиноген превращаются, соответственно, в стеркобилин и уробилин. Система комплемента Система комплемента — это сложный комплекс белков сыворотки крови, который является одним из компонентов неспецифического иммунитета и активируется по типу ферментативно-каскадной реакции, т.е. продукт предыдущей реакции играет роль катализатора следующей. Б. Активные фракции системы комплемента активируют фагоцитоз (опсонины - С3b и C5b). В. Активные фракции системы комплемента принимают участие в формировании воспалительной реакции. Действие их на тучные клетки вызывает дегрануляцию последних. Анафилотоксины действуют также на гладкие мышцы, вызывая их сокращение. Действуют они и на стенку сосуда: вызывают активацию эндотелия и повышение его проницаемости, что создает условия для экстравазации (выхода) из сосудистого русла жидкости и клеток крови в ходе развития воспалительной реакции. С3а выступает в роли иммуносупрессора (т.е. подавляет иммунный ответ). С5а является иммуностимулятором (т.е. усиливает иммунный ответ).
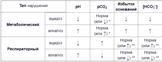
Билет №34 1)Витамин К (филлохинон, антигеморрагический) 4. Связывание ионов Са 2)Роль внутриклеточных посредников. 1. Мембранный – рецептор расположен на мембране. (пептидные и белковые гормоны, катехоламины, эйкозаноиды) 2. Цитозольный – рецептор расположен в цитозоле. Билет №35 1 ) Антиоксидантные витамины. ВИТАМИН Е (токоферол, витамин размножения) 2.Витамин А (каротиноид) — это сильный витамин антиоксидант. Благодаря наличию двойных связей в изопреновой цепи витамин осуществляет нейтрализацию свободных кислородных радикалов, особенно существенно эта функция проявляется у каротиноидов. 3. Витамин С Антиоксидантная роль: восстановление окисленного вита Е, лимитирование свободнорадик реакций в делящихся тк,ограничен воспаления,снижение ЛП в крови 2)Гормоны задней доли гипофиза.Гипофиз состоит из 3-х долей: передняя, средняя, задняя. Активируют: стресс,никотин, ангиотензин II,ацетилхолин,активация барорецепторов сердца и каротидного синуса возбуждение осморецепторов гипоталамуса и печени Уменьшают: этанол, глюкокортикоиды. Механизм действия Зависит от рецепторов: 1. Кальций-фосфолипидный механизм, сопряжен с V1-рецепторами гладких мышц артериол, печени, тромбоцитов,2. Аденилатциклазный механизм – с V2-рецепторами почечных канальцев. Мишени и эффекты ↑реабсорбцию воды в эпителиоцитах дистальных канальцев и собирательных трубочек,Поддерживает стабильное давление крови, стимулируя тонус сосудов: ↑ тонус гладких мышц сосудов кожи, скелетных мышц и миокарда (в меньшей степени),↑ чувствительность механорецепторов в каротидных синусах к изменениям артериального давления, памяти и поведенческих аспектах стресса, секрецию АКТГ и пролактина , Поддерживает минерализацию кости, усиливая активность как остеобластов, так и остеокластов. ↑вязкость крови, образ ф Виллебранда , ф VIIIи тканевого активатора плазминогена (t-PA),↑ глюкагона,↓липолитический эффект катехоламинов в жировой ткани, Патология Гипофункция несахарного диабета . Его частота примерно 0,5% всех эндокринных заболеваний. Проявляется большим объемом мочи до 8 л/сутки, жаждой и полидипсией, сухостью кожи и слизистых, вялостью, раздражительностью. Синдром неадекватной секреции – при образовании гормона какими-либо опухолями, при заболеваниях мозга. Окситоцин. Пептид. Стимулирует сокращение гладких мышц, в особенности матки, а также сокращение мышечных волокон вокруг альвеол молочных желез, вызывая секрецию молока. Это происходит благодаря проникновению ионов К+ в мышечные клетки и угнетению ацетилхолинэстеразы. Синтезируется паравентрикулярным ядром гипоталамуса. Функции: стим. секрецию молока; стим. сокращение матки; рилизинг фактор для выброса пролактина. 3)Регуляция и поддержание кислотно-основного равновесия. КОС – активность физико-химических процессов, составляющих функционально единую систему стабилизации концентрации ионов Н+. Основ характер= (рН) Нарушение 1. Накопление кислых веществ:(↑ поступление кислых продуктов, ↓ удаление кислых продуктов. ) 2.Недостаток кислых веществ:(избыточное удаление угольной кислоты) 3. Накопление щелочных веществ:↑поступление, 4. Недостаток щелочных в-в: потери бикарбонат-ионов, Диагностика 1.Определение рН . ↓Ацидоз,↑алкалоз,компенсаторный (норм ph)2.Определение величины рСО 2.3.Сопоставление
Показатели: 1) ph=7,35-7,45 2) рСО2(+ концентрацию СО2, растворенного в плазме крови) H2CO3 и HCO3‑.=35-45 мм рт. ст.3) рО2 =.83-108 мм рт. ст.4) [HCO3‑]Концентрация бикарбонат-ионов в плазме крови. Критер.метабол.наруш =21-24 ммоль/л5)ВВПоказывает концентрацию всех буферных оснований в цельной крови.43,7-53,5 ммоль/л6)BEИзбыток оснований. Показывает колебания количества основных эквивалентов. «+»= -↓ Н+. «-«=↑Н+. 0 ± 2,5 ммоль/л 7) Регуляция.Система химических реакций: действие вне- и внутриклеточных буферных систем (бикарбонатная, фосфатная, белковая, гемоглобиновая), интенсивность внутриклеточного образования ионов Н+ и НСО3–. 2. Система физиологических механизмов: 1)легочная вентиляция и удаление СО2. 2)почечная (Реабсорбция бикарбонатных ионов HCO3-Ацидогенез – удаление ионов Н+ с титруемыми кислотами Аммониегенез – удаление ионов Н+ в составе ионов аммония NH4+) Буферные системы – это соединения, противодействующие резким изменениям концентрации ионов Н Бикарбонатная буферная система –(10%). Бикарбонатная система представляет собой сопряженную КО пару, Н2СО3, выполняющую роль донора протона, и НСО3–, выполняющего роль акцептора протона. ↑ Н+ + НСО3–, => Н2СО3. Далее ↓Н2СО3 при выделении СО2 через легкие в результате их гипервентиляции Фосфатная буферная система( 2%) от крови и до 50% мочи. Она образована гидрофосфатом (HPO42–) и дигидрофосфатом (H2PO4–). Первое соединение слабо диссоциирует и ведет себя как слабая кислота, второе обладает щелочными свойствами. В норме отношение HРO42– к H2РO4– равно 4 : 1. Н++ HPO42‑= H2PO4–=>↓ Н+ При ↑ОН–‑групп нейтр от H2PO4–ионами Н+: Белковый буфер. альбумин, иблагодаря амфотерным свойствам. ( 5%.) В кислой среде ↓ диссоциация СООН‑групп аминокислотных радикалов , а группы NH2 +Н+ =>белок заряд +. В щелочной среде ↑диссоциация COOH‑групп, Н++ ↑ОН–‑ионов = pH сохраняется. Белки заряд «-« Гемоглобиновый буфер 28%. Самый мощный. Кислота H‑HbO2. Основание - Н‑Нb. Тесно работает с бикарбонатной бф сис. Изменение кисл гемоглобина в тка и в легких=>связыв H+ или О2. Механизм буфера + или - H+остатком гистидина в глобиновой части молекулы (эффект Бора).В тканях более кислый Ph. Ионы H+ + (HbО2) и =H‑HbО2. => отчада О2 (эффект Бора) => H‑Hb. НbO2+ Н+ → [H-HbO2] → Н-Hb + O2 В результате↓ Н2СО3, продуцируются ионы НСО3‑ и тканевое пространство подщелачивается.В легких после удаления СО2 происходит защелачивание крови. При этом +О2 к H-Hb = > кислоту более сильную, чем угольная. Она отдает свои ионы Н+ в среду, предотвращая ↑Рн. Н-Hb + O2 → [H-HbO2] → НbO2 + Н+
Билет №36 1) Гормональная регуляция обмена кальция и роль витамина D За обмен кальция и фосфатов в организме отвечают три гормона – кальцитриол, кальцитонин и паратиреоидный гормон. Кальцитриол -производное витамина D и относится к стероидам. Синтез В коже под действием УФ и поступающие с пищей холекальциферол (витамин D3) и эргокальциферол (витамин D2) гидроксилируются в гепатоцитах по С25 и в эпителии проксимальных канальцев почек по С1. В результате формируется 1,25-диоксихолекальциферол (к альцитриол). Активируют : Гипокальциемия ↑гидроксилирование витамина D 1 в почках через ↑секреции паратгормона, стимулирующего этот процесс. ↓: Избыток кальцитриола подавляет гидроксилирование в почках. Мишени и эффекты Эффекты регуляции концентрации кальция и фосфора в крови: в кишечнике индуцирует синтез белков, отвечающих за всасывание кальция и фосфатов, в почечных канальцах ↑ синтез белков-каналов для реабсорбции ионов кальция и фосфатов, в костной ткани усиливает активность остеобластов и остеокластов, способствует минерализации кости остеобластами,и участвует в проведении нервного возбуждения ,,поддерживает тонус сердечной и скелетной мускулатуры , и влияет на пролиферацию клеток. 3. Также действие кальцитриола сопровождается подавлением секреции п аратиреоидного гормона. Гипофункция -Рахит Мишени и эффекты ↑ концентрации кальция и ↓ концентрации фосфора в крови.Актив остеокласты и происходит деструкция костной ткани, при ↓концентрациях активируется перестройка кости и остеогенез.↑реабсорбция кальция и магния,↓реабсорбция фосфатов, аминокислот, карбонатов, натрия, хлоридов, сульфатов. образование кальцитриола,при участии кальцитриола усиливается всасывание кальция и фосфатов. Гипофункция Возникает при случайном удалении железы при операциях на щитовидной железе, аутоиммунной деструкции ткани желез. Возникающая гипокальциемия и гиперфосфатемия проявляется в виде высокой нервно-мышечной возбудимости, судорог, тетании. При резком снижении кальция возникает дыхательный паралич, ларингоспазм. Гиперфункция Первичный гиперпаратиреоз возникает при аденоме желез. Нарастающая гиперкальциемия вызывает повреждение почек, мочекаменную болезнь. Вторичный гиперпаратиреоз является результатом почечной недостаточности, при которой происходит нарушение образования кальцитриола, ↓концентрации кальция и компенсаторное ↑ синтеза паратиреоидного гормона. Кальцитонин Механизм действия Аденилатциклазный Мишени и эффекты Эффект кальцитонина заключается в ↓ концентрации кальция и фосфора в крови:в костной ткани подавляет активность остеокластов, что улучшает вход кальция и фосфатов в кость,в почках подавляет реабсорбцию ионов Ca2+, фосфатов, Na+, K+, Mg2+. 2) Регуляция и поддержание кислотно-основного равновесия. КОС – активность физико-химических процессов, составляющих функционально единую систему стабилизации концентрации ионов Н+. Основ характер= (рН) Нарушение 1. Накопление кислых веществ:(↑ поступление кислых продуктов, ↓ удаление кислых продуктов. ) 2.Недостаток кислых веществ:(избыточное удаление угольной кислоты) 3. Накопление щелочных веществ:↑поступление, 4. Недостаток щелочных в-в: потери бикарбонат-ионов, Диагностика 1.Определение рН . ↓Ацидоз,↑алкалоз,компенсаторный (норм ph)2.Определение величины рСО 2.3.Сопоставление
Показатели: 1) ph=7,35-7,45 2) рСО2(+ концентрацию СО2, растворенного в плазме крови) H2CO3 и HCO3‑.=35-45 мм рт. ст.3) рО2 =.83-108 мм рт. ст.4) [HCO3‑]Концентрация бикарбонат-ионов в плазме крови. Критер.метабол.наруш =21-24 ммоль/л5)ВВПоказывает концентрацию всех буферных оснований в цельной крови.43,7-53,5 ммоль/л6)BEИзбыток оснований. Показывает колебания количества основных эквивалентов. «+»= -↓ Н+. «-«=↑Н+. 0 ± 2,5 ммоль/л 7) Регуляция.Система химических реакций: действие вне- и внутриклеточных буферных систем (бикарбонатная, фосфатная, белковая, гемоглобиновая), интенсивность внутриклеточного образования ионов Н+ и НСО3–. 2. Система физиологических механизмов: 1)легочная вентиляция и удаление СО2. 2)почечная (Реабсорбция бикарбонатных ионов HCO3-Ацидогенез – удаление ионов Н+ с титруемыми кислотами Аммониегенез – удаление ионов Н+ в составе ионов аммония NH4+) Буферные системы – это соединения, противодействующие резким изменениям концентрации ионов Н Бикарбонатная буферная система –(10%). Бикарбонатная система представляет собой сопряженную КО пару, Н2СО3, выполняющую роль донора протона, и НСО3–, выполняющего роль акцептора протона. ↑ Н+ + НСО3–, => Н2СО3. Далее ↓Н2СО3 при выделении СО2 через легкие в результате их гипервентиляции Фосфатная буферная система( 2%) от крови и до 50% мочи. Она образована гидрофосфатом (HPO42–) и дигидрофосфатом (H2PO4–). Первое соединение слабо диссоциирует и ведет себя как слабая кислота, второе обладает щелочными свойствами. В норме отношение HРO42– к H2РO4– равно 4 : 1. Н++ HPO42‑= H2PO4–=>↓ Н+ При ↑ОН–‑групп нейтр от H2PO4–ионами Н+: Белковый буфер. альбумин, иблагодаря амфотерным свойствам. ( 5%.) В кислой среде ↓ диссоциация СООН‑групп аминокислотных радикалов , а группы NH2 +Н+ =>белок заряд +. В щелочной среде ↑диссоциация COOH‑групп, Н++ ↑ОН–‑ионов = pH сохраняется. Белки заряд «-« Гемоглобиновый буфер 28%. Самый мощный. Кислота H‑HbO2. Основание - Н‑Нb. Тесно работает с бикарбонатной бф сис.Изменение кисл гемоглобина в тка и в легких=>связыв H+ или О2. Механизм буфера + или - H+остатком гистидина в глобиновой части молекулы (эффект Бора).В тканях более кислый Ph. Ионы H+ + (HbО2) и =H‑HbО2. => отчада О2 (эффект Бора) => H‑Hb.В результате↓ Н2СО3, продуцируются ионы НСО3‑ и тканевое пространство подщелачивается.В легких после удаления СО2 происходит защелачивание крови. При этом +О2 к H-Hb = > кислоту более сильную, чем угольная. Она отдает свои ионы Н+ в среду, предотвращая ↑Рн. Н-Hb + O2 → [H-HbO2] → НbO2 + Н+ 3) Минералокортикоиды. Дезоксикортикостерон и альдостерон(биосинтез в клубочковой зоне надпочечников) Образованный из холестерола прогестерон на пути к альдостерону подвергается последовательному окислению 21-гидроксилазой, 11-гидроксилазой и 18-гидроксилазой. В конечном итоге образуется альдостерон.Регуляция реабсорбции почечными канальцами Na и активация секреции К. регулируют обмен Na, K, CL и воды. Удерживают ионы Na и CL в организме и выводят с мочой ионы K. Период полураспада кортикостероидов составляет всего 70-90 мин. Продукты окисления этих гормонов – 17 –кетостероиды выводятся с мочой. Определение 17 –кетостероидов в моче имеет большое клиническое значение. ↑ их экскреция может быть при опухолях надпочечников. Гормоны надпочечников широко используются в клинической практике. Цитозольный. Мишени и эффекты Воздействует на слюнные железы, на дистальные канальцы и собирательные трубочки почек. В почках усиливает реабсорбцию ионов Na и потерю ионов калия посредством следующих эффектов: ↑Na+,K+-АТФазы на базальной мембране эпителиальных клеток,стимулирует синтез митохондриальных белков и увеличение количества нарабатываемой в клетке энергии для работы Na+,K+-АТФазы,стимулирует образование Na-каналов на апикальной мембране клеток почечного эпителия.
Билет№1 Предмет и задачи биохимии(кафедре 110 лет) БХ-наука о хим.составе и св-вах живых орг. в процессе жизнедеятельности.(+обмен в-тв). 2)»Ключевые» ферменты, их роль в обмене веществ . Присоединение эффектора к аллостерической (регуляторной) субъединице изменяет конформацию белка и, соответственно, активность каталитической субъединицы. Аллостерические ферменты обычно стоят в начале метаболических путей, и от их активности зависит течение многих последующих реакций. Поэтому они часто называются ключевыми ферментами. В качестве отрицательного регулятора может выступать конечный метаболит биохимического процесса или продукт данной реакции, т.е включается механизм обратной отрицательной связи. Если регуляторами являются начальный метаболит или субстрат реакции, то говорят о прямой регуляции, она может быть как положительной, так и отрицательной. Также регулятором могут быть метаболиты биохимических путей, каким то образом связанных с данной реакцией. Например: гликолиз:- Гексокиназа—аллостерически ингибируется глюкозо-6-фосфатом. регуля фермент в гепатоцитах. Синтез глюкокиназы индуцируется инсулином. 2. Фосфофруктокиназа-1. катализирует реак-ю, лимит скорость всего процесса (наиболее медленная реакция). индуцируется инсулином. активаторы — АМФ, АДФ, фруктозо-2,6- дифосфат. Уровень фруктозо-2,6-дифосфата увеличивается под действием инсулина и понижается под действием глюкагона. Аллостерические ингибиторы — АТФ, цитрат. 3. Пируваткиназа.Фермент активен в нефосфорилированной форме. Глюкагон (в гепатоцитах) и адреналин (в миоцитах) инактивируют фермент. Инсулин активирует. Аллостерический активатор — Фр-1,6-ФФ. Аллостерический ингибитор — АТФ, ацетил КоА. ц.кребса: цитратсинтаза (1-я реакция) 2) изоцитратдегидрогеназа (3-я реакция) аллостерически ингибируются избытком АТФ и НАДН2. Изоцитратдегидрогеназа ингибируется АТФ намного сильнее, чем цитратсинтаза, поэтому в условиях энергетического покоя повышается концентрация цитрата, и он выходит в цитоплазму по градиенту концентраций путем облегченной диффузии. В цитоплазме цитрат превращается в Ацетил-КоА, который участвует в синтезе жирных кислот. Пируватдегидрогеназный комплекс: -пируватдегидрогеназа -дегидролипоилацетилтрансфераза -дегидролипоилдегидрогеназа Пентозо-фосфорн путь: глюкозо-6-фосфатдегидрогеназа и дегидрогеназа 6-фосфоглюконата. Индукторами -инсулин. Активность ↑при углеводов в организм и ↓голодании и диабете. 3)Механизм действия гормонов белковой природы.Инсулин. Гормоны (греч. hormao – привожу в движение) – это в-ва, вырабатываемые спец кл и регул обмен в-в в отдельных органах и в организме. большая специфичность действия и ↑биологическая активность. Синтез – препроинсулина-проинсулина-инсулина. Рецептор: тирозинкиназная система.На мембр Очень быстрые (секунды) - гиперполяризация мембран клеток, изменение мембранного транспорта глюкозы, ионов.Быстрые (минуты) - активация ферментов, что приводит анаболизму.Медленные (минуты - часы) - ↑потребления клеткой аминокислот, избирательная индукция ферментов анабализма или репрессия ферментов катаболизма (действие на геном). Самые медленные (часы - сутки) - митоз, размножение клеток. Билет №2 1)Классификация белков. Структуры белков и методы их определения. По химическому сост.1) Простые белки (протеины) гидролизуются кислотами или щелочами до аминокислот и не дают при гидролизе других органических и неорганических соединений. 1. Альбумины –б плазмы крови с массой40 кДа ( кислые .↑ отрицательный заряд) транспортер для билирубина и жирных кислот. 2.Глобулины – белки, слаб кис. Нейтр. Белок плазмы крови.3. Проламины – белки, растворяются в 60-80%-ном этилового спирта. Пролина. Раст происход 4.Глютелины – белки, которые извлекаются из растительных тканей разбавленными растворами щелочей (0,2%-ный NаОН). 5. Протамины – это белки ↓массы, состоящие на 80% основ св-ва. Аргинин.6. Гистоны –↓белки также основного характера, содержатся в хромосомах клеточных ядер и играют важную роль в образовании структуры хроматина. 2) Сложные белки содерж простетическую группу. 1.Хромопротеины(гемопротеины,магнийпорфирины,флавопротеины) 2.гемопротеины (белок+гем) гем и его произвдные.3 флавопротеины (белок+изоаллоксазиновые производные ФМН и ФАД). Фермент катал ОВР 4.магнийпорфирины 5 нуклеопротеины (белок+нк к-та)6. Липопротеины 7гликопротеины (бел+углевод) 8 металлопротеины (б+ме) фосфопротеины (б+фосфат( 2)Переваривание и всасывание углеводов в ЖКТ Переваривание углеводов |
Последнее изменение этой страницы: 2019-04-11; Просмотров: 329; Нарушение авторского права страницы




 .
.  Аминогруппы разных аминокислот в ходе реакций трансаминирования переносятся на пируват, источником которого служат глюкоза и безазотистые остатки аминокислот. Особенно много аланина выделяют мышцы в силу их большой массы, а также потому, что работающие мышцы часть энергии получают за счет распада аминокислот.Аланин →печень,
Аминогруппы разных аминокислот в ходе реакций трансаминирования переносятся на пируват, источником которого служат глюкоза и безазотистые остатки аминокислот. Особенно много аланина выделяют мышцы в силу их большой массы, а также потому, что работающие мышцы часть энергии получают за счет распада аминокислот.Аланин →печень,

 Выделяют три основных решения уравнения Михаэлиса-Ментен:
Выделяют три основных решения уравнения Михаэлиса-Ментен:


 2) Современное представление о биологическом окисление. Типы биолог окисления. Микросомальное окисление
2) Современное представление о биологическом окисление. Типы биолог окисления. Микросомальное окисление