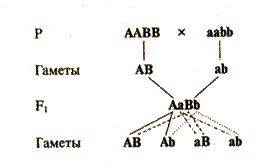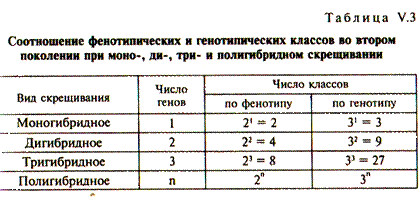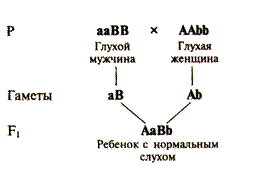|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
V.5. ПОЛИГИБРИДНЫЕ СКРЕЩИВАНИЯ
Основные закономерности, открытые Г.Менделем, касались наследования и расщепления только по одной паре альтернативных признаков (при моногибридном скрещивании). На следующем этапе Менделя интересовал вопрос, какими признаками будет обладать потомство от скрещивания родительских форм, различающихся одновременно несколькими признаками. Гибриды, полученные от скрещивания особей, различающихся одновременно по двум парам альтернативных признаков, носят название дигибридов.Рассмотрим результаты классического опыта Менделя по дигибридному скрещиванию. Для скрещивания были отобраны растения, которые имели гладкие желтые горошины (оба признака доминантные), и растения с морщинистыми зелеными горошинами (оба признака рецессивные). Согласно первому закону все потомство было единообразно: дигибридные растения давали только гладкие желтые семена. В потомстве от самоопыления 15 дигибридных растений было получено 556 горошин: из них 315 гладких желтых, 108 гладких зеленых, 101 морщинистых желтых и 32 горошины были морщинистые зеленые. Это соотношение близко к соотношению 9: 3: 3: 1 иотражает относительные частоты 4 классов фенотипов. При анализе по каждому признаку в отдельности (только по форме или только по окраске) расщепление гибридов в F2 соответствовало соотношению 3: 1. Проследим расщепление по признаку формы семян. Гладких горошин было 315 + 108, что составило в сумме 423 гладких. Морщинистых горошин было 101 + 32, что составило 133 семян. Отношение 423 к 133 было близко к уже известному отношению 3: 1. То же самое отношение было получено при анализе расщепления по признаку окраски. Желтых горошин вне зависимости от их формы было 416 (315 + 101), а количество зеленых составило 140 горошин (108 + 32). Отношение также близко к 3: 1. Полученные результаты свидетельствовали, что расщепление по каждой паре аллелей при дигибридном скрещивании происходит как два независимых события. Таким образом, соотношение фенотипов при дигибридном скрещивании представляет собой результат случайного или независимого объединения результатов двух моногибридных скрещиваний. Этот вывод отражает сущность третьего закона Менделя — закона независимого комбинированиягенов. Обозначив доминантный аллель, определяющий желтую окраску, буквой А и рецессивный аллель, определяющий зеленую окраску, буквой а, буквой В — доминантный аллель, контролирующий развитие гладкой формы горошины, и соответственно рецессивный аллель, контролирующий развитие морщинистой формы, буквой b, дигибридное скрещивание можно записать следующим образом (рис. V.4). Для того чтобы представить фенотипические и генотипические классы потомков дигибридов первого поколения, воспользуемся так называемой решеткой Пеннета, которая позволяет установить все возможные сочетания мужских и женских гамет (табл. V.2). Легко убедиться, что 9 из 16 (т.е. 9/16) теоретически ожидаемых потомков имеют одновременно два доминантных признака (желтые гладкие горошины — жг); 3/16 — доминантный и рецессивный признак (желтые морщинистые — жм); 3/16 — рецессивный и доминантный признаки (зеленые гладкие — зг) и l/l6 часть потомков имеет одновременно два рецессивных признака (зеленые и морщинистые — зм). При анализе генотипов по решетке Пеннета мы обнаруживаем 9 различных классов в соотношении 1: 2: 2: 4: 1: 2: 1: 2: 1. Расщепление в дигибридном скрещивании в F2 по фенотипу и генотипу можно получить, перемножая относительные частоты отдельных фенотипов или отдельных генотипов, поскольку гены, контролирующие развитие различных признаков, наследовались независимо друг от друга. По каждому из независимых признаков (окраска горошин и характеристика их поверхности) отношение частот, как было ранее показано, составляет 3: 1. Тогда, перемножая (Зж: 1з) на (Зг: 1м), получаем 9жг: 3жм: 3зг: 1зм, что точно соответствует данным решетки Пеннета.
Рис. V.4. Схема дигибридного скрещивания. Гибриды первого поколения единообразны как по фенотипу, так и по генотипу. Они образуют гаметы четырех различных типов — АВ, Ab, аВ, ab
Зная, что при моногибридном скрещивании расщепление по генотипу соответствует 1АА: 2Аа: 1аа для одной пары и 1BB: 2Bb: lbb для другой, можно подсчитать частоты, или вероятности, генотипов различных классов. Вероятности генотипов соответствуют: АА - 1/4, Аа - 1/2, аа - 1/4; ВВ - 1/4, Вb - 1/2, bb - 1/4. Например, относительная частота генотипа ААВВ рассчитывается путем перемножения вероятностей 1/4АА x 1/4ВВ = 1/16ААВВ, для ААВb — 1/4АА х 1/2 Вb = 1/8 или 2/l6, ААВЬ. Тем же путем получаем распределение всех остальных различающихся по генетической конституции классов особей в отношении 1: 2: 2: 4: 1: 2: 1: 2: 1, что также полностью соответствует данным решетки Пеннета. Поступая аналогичным образом, можно представить результаты расщепления по фенотипу и генотипу для тригибридного скрещивания, когда родительские формы различаются по трем независимым признакам и в F1 образуются тригибриды. Эксперименты показывают, что при тригибридном скрещивании расщепление в F2 по фенотипу дает 8 различных классов особей в соотношении 27: 9: 9: 9: 3: 3: 3: 1, а расщепление по генотипу дает 27 раз-, личных классов. Подобным образом возможен расчет вероятностей фенотипических и генотипических классов для любого пол и гибридного скрещивания (табл. V.3). В обoем виде эти соотношения можно выразить простыми формулами: число фенотипических классов равно 2n, где «2» отражает парность аллелей, а показатель степени «n» — число независимых генов. Число генотипических классов равно Зn, где основание степени «3» — число генотипических классов при моногибридном скрещивании, а показатель степени «n» — число генов. Очевидно, что в основе приведенных формул лежат закономерности моногибридного скрещивания. Они справедливы для любого числа генов, но не превышающих гаплоидное число п. Важно отметить, что закономерности, открытые Менделем, реализуются при анализе большого количества особей, поскольку малое количество в потомстве гибридов (например, дети одной семьи) может давать отклонения от точного соотношения ожидаемых классов расщепления в силу случайных событий. Гибридологический анализ, разработанный Менделем, и результаты, полученные на его основе, заложили концепцию фундаментального понятия генетики и биологии в целом — понятие гена. В последние десятилетия XIX в. были обнаружены хромосомы, описаны митотическое и мейотическое деления клетки. Тем не менее не были известны материальные носители наследственной информации. Только после того как законы Менделя были открыты вновь в 1900 г., сопоставление менделевского расщепления признаков и распределения хромосом в мейозе позволило сделать окончательный вывод о том, что именно хромосомы являются носителями генетической информации. Этими событиями ознаменовалось начало нового научного периода развития генетики, а наблюдения и выводы Менделя и в настоящее время составляют важнейшую главу учения о наследственности и изменчивости.
V.6. ВЗАИМОДЕЙСТВИЕ ГЕНОВ
В предыдущем разделе мы рассматривали закономерности межаллельных взаимоотношений, т.е. взаимоотношений между аллелями одного и того же гена: доминирование, неполное доминирование, кодоминирование, серию множественных аллелей. Однако большое количество признаков формируется при участии нескольких генов, взаимодействие которых существенным образом отражается на особенностях фенотипа и приводит к отклонению от менделевской закономерности расщепления фенотипов. Описано несколько типов взаимодействия неаллельных генов: комплементарность, эпистаз и полимерия. Комплементарное ib — взаимодействие неаллельных генов, которые обусловливают развитие нового признака, отсутствующего у родителей. Примером комплементарного действия у человека могут служить случаи, когда у глухих родителей рождаются дети с нормальным слухом. Развитие нормального слуха находится под генетическим контролем десятков различных неаллельных генов, гомозиготное рецессивное состояние одного из которых может приводить к одной из форм наследственной глухоты. Таких форм у человека известно более 30. Если один из родителей является гомозиготой по рецессивному гену аа (рис. V.5), а другой — гомозиготой по другому рецессивному гену bb, то все их дети будут двойными гетерозиготами и, следовательно, слышащими, поскольку доминантные аллели будут взаимно дополнять друг друга (см. рис. V.5). Таким образом, формируется новый по отношению к родителям признак — нормальный слух.
Рис. V.5. Схема, поясняющая возможность рождения детей с нормальным слухом у глухих родителей с различными генетическими формами глухоты
Эпистаз (от греч. epi — над + stasis — препятствие) — взаимодействие неаллельных генов, при котором наблюдается подавление проявления одного гена действием другого, неаллельного гена. Подавляющий ген называется геном-супрессором, а подавляемый — гипостатическим геном. По-видимому, действие гена-супрессора на подавляемый ген сходно с принципом доминантность — рецессивность. Но существенное различие заключается в том, что эти гены не являются аллельными, т.е. расположены в негомологичных хромосомах или занимают различные локусы в гомологичных. Различают доминантный и рецессивный эпистаз. При доминантном эпистазе доминантный аллель гена-супрессора подавляет проявление доминантного аллеля другого гипостатического гена. При рецессивном эпистазе, или криптомерии, рецессивный аллель гена-супрессора, будучи в гомозиготном состоянии, не дает проявиться доминантной или рецессивным аллелям других генов. Примером рецессивного эпистаза у человека может служить так называемый бомбейский фенотип, когда индивид, имеющий доминантный аллель группы крови системы АВО (например, аллель В, определяющий принадлежность человека к IIIили IV группе), идентифицируется в реакции агглютинации как человек с 0(1). Это состояние возникает в результате того, что данный индивид является рецессивной гомозиготой (hh) по другой, нежели система АВО, генетической системе Hh. Для реализации аллелей 1А и IB необходимо присутствие хотя бы одного доминантного аллеля Н. Полимерия (от греч. polys — много + meros — часть) — вид взаимодействия, когда эффекты нескольких неаллельных генов, определяющих один и тот же признак, примерно одинаковы. Подобные признаки получили название количественных, или полимерных признаков. Как правило, степень проявления полимерных признаков зависит от числа доминантных генов. Наследование полимерных признаков было впервые описано шведским генетиком Г. Нильсон-Эле в 1908 г. Проводя скрещивание различных форм пшеницы (с красными и белыми зернами), он наблюдал расщепление в F2 признака окраски в отношении: 15/l6 окрашенных и 1/l6 белых. Среди окрашенных зерен он наблюдал все переходы — от интенсивно окрашенных до слабо окрашенных. Анализ особенностей расщепления показал, что в данном случае окраску зерен определяют два доминантных аллеля двух различных генов, а сочетания их рецессивных аллелей определяют отсутствие окраски. Поскольку полимерные гены имеют однонаправленное действие, их, как правило, обозначают одинаковыми буквами. Таким образом, исходные родительские формы имели генотипы A1A1A2A2 и a1a1a2a2. Наличие всех четырех доминантных аллелей определяло самую интенсивную окраску, трех доминантных аллелей (типа А1А1А2а2) — менее интенсивную окраску и т.д. Примером полимерного наследования у человека является наследование окраски кожных покровов. В браке индивида негроидной расы (коренного жителя Африки) с черной окраской кожи и представителем европеоидной расы с белой кожей дети рождаются с промежуточным цветом кожи (мулаты). В браке двух мулатов потомки могут обладать любой окраской кожи: от черной до белой, поскольку пигментация кожи обусловлена действием трех или четырех неаллельных генов. Влияние каждого из этих генов на окраску кожи примерно одинаково. Полимерное наследование характерно для так называемых количественных признаков, таких, как рост, вес, окраска кожных покровов, скорость протекания биохимических реакций, артериальное давление, содержание сахара в крови, особенности нервной системы, уровень интеллекта, и многих других, которые нельзя разложить на четкие фенотипические классы. Чем большее число неаллельных генов контролируют развитие количественного признака, тем менее заметны переходы между фенотипическими классами.
Популярное: |
Последнее изменение этой страницы: 2016-03-25; Просмотров: 1091; Нарушение авторского права страницы