|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
IV.3. СОХРАНЕНИЕ ИНФОРМАЦИИ ОТ ПОКОЛЕНИЯ К ПОКОЛЕНИЮ
При размножении зигота, образовавшаяся в результате слияния гамет, дает начало миллионам и миллиардам клеток тела. Каждая исходная молекула ДНК дает начало двум новым молекулам ДНК, с сохранением в неизменном виде всех особенностей исходной молекулы. Процесс удвоения ДНК, происходящий во время синтетической (S) стадии интерфазы, носит название репликации. Во время репликации информация, закодированная в последовательности нуклеиновых оснований молекулы родительской ДНК, передается с максимальной точностью дочерним ДНК. Вкратце рассмотрим схему репликации. В 1956 г. А.Корнберг выделил фермент, который был способен связывать свободные нуклеотиды друг с другом, и дал ему название ДНК-полимераза. Впоследующие годы были обнаружены идругие виды полимераз, также способных удлинять цепи ДНК в направлении 5'→ 3', последовательно прибавляя по одному нуклеотиду к З'-ОН-концу цепи. ДНК-полимераза может строить лишь одну цепь молекулы ДНК. Обнаружение ДНК-полимеразы стало первым практическим шагом анализа биохимических механизмов репликации. Способ репликации, характерный для всех эукариот, и том числе и человека, известен под названием полуконсервативной репликации (рис. IV. 10). Вначале процесса репликации особый фермент — хеликаза (от греч. helix — спираль) — расплетает родительскую ДНК на две нити, каждая из которых служит матрицей, определяющей последовательность новой, комплементарной цепи ДНК. При полуконсервативной репликации дочерние клетки первого поколения получают только одну из нитей ДНК родительской клетки. Вторая нить синтезируется заново, при этом она комплементарна родительской цепи. Процесс повторяется при образовании дочерних клеток второго поколения. Таким образом, только две из четырех дочерних клеток второго поколения содержат по одной цепи исходной родительской ДНК.
Рис. IV. 10. Схема полуконсервативной репликации ДНК
Поскольку ДНК-полимераза катализирует репликацию только в одном направлении (5'→ 3'), непрерывно достраивается только одна новая цепь молекулы ДНК (смысловая). Вторая цепь (антисмысловая) синтезируется другой ДНК-полимеразой, движущейся в обратном направлении, в виде коротких участков ДНК. Затем эти фрагменты ДНК связываются в единую цепь ферментом ДНК-лигазой. Таким образом, репликация ДНК обеспечивает высочайшую точность воспроизведения генетической информации, заключенной в последовательности оснований ДНК и тем самым реализует основные функции ДНК — сохранение генетической информации и точное ее воспроизведение в ряду поколений. Сразу после открытия структуры двойной спирали ДНК стало очевидно, что сама молекула ДНК не может служить матрицей для синтеза белка. Этот вывод логично вытекал из того, что почти вся ДНКнаходится в хромосомах, расположенных в ядре, в то время как синтез белка осуществляется в цитоплазме клетки. Таким образом, генетическая информация, зашифрованная в структуре ДНК, должна передаваться иной молекуле, способной транспортироваться в цитоплазму и участвовать в синтезе белка. Такой молекулой оказалась мРНК, а процесс образования мРНК получил название транскрипции. Транскрипция — это процесс переноса генетической информации от ДНК к РНК. Все виды РНК (мРНК, тРНК, рРНК и гяРНК) синтезируются в соответствии с последовательностью нуклеотидов в молекуле ДНК, служащей для них матрицей. Процесс транскрипции осуществляется при участии трех ДНК-зависимых РНК-полимераз. Полимераза I находится в ядрышках, где катализирует синтез рРНК. Полимеразы II и IIIнаходятся в кариоплазме, где полимераза И контролирует синтез первичного транскрипта мРНК, а полимераза IIIучаствует в синтезе тРНК. Процесс транскрипции осуществляется следующим образом. РНК-полимераза, прикрепляясь к началу участка ДНК, расплетает двойную спираль. Перемещаясь вдоль одной из нитей, она последовательно выстраивает комплементарную нить РНК. По мере передвижения РНК-полимеразы растущая нить РНК отходит от матрицы ДНК, и двойная спираль ДНК позади фермента восстанавливается. Когда РНК-полимераза достигает специфического участка ДНК, называемого терминационным, рост цепи РНК прекращается, и происходит отделение ее от ДНК. Синтезированная нить РНК содержит информацию, точно переписанную с соответствующего участка ДНК. Процесс транскрипции, как и репликации, осуществляется при строгом соблюдении правила комплементарности с одним изменением: напротив аденина молекулы ДНК при транскрипции в молекулу РНК встраивается урацил (рис. IV. 11).
Рис. IV.11. Реализация правила комплементарности при матричном синтезе РНК
После того как заканчивается транскрипция, все виды РНК претерпевают определенные изменения, в результате которых они приобретают возможность выполнять специфические для каждойиз них функции. Подобное созревание РНК носит название процессинга. Матричная, или информационная, РНК (мРНК). На долю мРНК приходится примерно 3 — 5% всей содержащейся в клетке РНК. Молекулы мРНК образуются из больших молекул-предшественников — гяРНК. Изменения гяРНК связаны с модификацией по 5'- и З'-концам и сплайсингом. Изменение 5'-конца приводит к образованию специфической последовательности, называемой кэп-структурой. Изменение З'-конца заключается в присоединении к нему 120—150 остатков аденина (poly А). Сплайсинг — это процесс удаления участков молекулы РНК, соответствующих интронным последовательностям ДНК. Зрелая мРНК выходит в цитоплазму и становится матрицей для синтеза белка, т.е. переносит информацию о структуре белка от ДНК к рибосомам. Рибосомальная РНК (рРНК) составляет более 80% всей РНК клетки. Она кодируется генами, расположенными в так называемых ядрышковых организаторах некоторых акроцентрических хромосом. Последовательность нуклеотидов в рРНК сходна у всех организмов. Вся рРНК находится в цитоплазме, где она образует сложный комплекс с белками, формируя рибосому. На рибосомах информация, зашифрованная в структуре мРНК, переводится (транслируется) в аминокислотную последовательность, т.е. происходит синтез белка. На долю транспортных РНК (тРНК) приходится примерно 15% всей клеточной РНК. Поскольку большинство аминокислот кодируются несколькими триплетами, число различных тРНК значительно больше числа аминокислот (больше 20). Все молекулы тРНК имеют сходную структуру, напоминающую клеверный лист (рис. IV. 12). На 5'-конце молекулы всегда находится гуанин, а на 3'-конце — последовательность ЦЦА; тРНК узнает соответствующий кодон в мРНК ипереносит нужную аминокислоту в растущей полипептидной цепи. Узнавание кодона мРНК осуществляется с помощью антикодона транспортной РНК — специфичной для каждой аминокислоты последовательности трех оснований тРНК, комплементарных данному кодону мРНК. Аминокислота присоединяется к З'-концу тРНК с помощью фермента аминоацил-тРНК-синтетазы. Таким образом, тРНК играют связующую роль между мРНК и белком.
Рис. IV. 12. Строение транспортной РНК
IV.4. ТРАНСЛЯЦИЯ
Трансляция («перевод») — это процесс реализации информации, закодированной в структуре мРНК, в последовательность аминокислотных остатков белка. Центральное место в трансляции принадлежит рибосомам, в большом количестве присутствующим в цитоплазме клеток. Функция рибосомы состоит в том, чтобы удержать в нужном положении мРНК, тРНК и белковые факторы до тех пор, пока не образуется пептидная (белковая) связь между соседними аминокислотами. Рибосома образована двумя субъединицами — большой и малой, состоящими из рибосомальной РНК (рРНК) и белков. Аминокислоты, находящиеся в цитоплазме, доставляются в рибосомы посредством транспортных РНК (тРНК); мРНК содержат транскрибированную с соответствующих генов информацию о том, какие белки должны быть синтезированы в рибосомах, и служат матрицами, определяющими, в какой последовательности аминокислоты, доставленные тРНК в рибосомы, включаются в растушую полипептидную цепь белка. В области большой субъединицы рибосомы между аминокислотами образуются пептидные связи, т. е. происходит считывание информации. Как только новая аминокислота присоединилась к растущей цепи, рибосома перемещается по нити мРНК и узнает следующий кодон. Молекула тРНК, освободившись от аминокислоты, уже включенной в полипептидную цепь, покидает рибосому и возвращается в цитоплазму, чтобы вновь образовать комплекс с аминокислотой. Синтез белковой цепи продолжается до тех пор, пока процесс не доходит до одного из стоп-кодонов: УАА, УАГ или УГА. Процесс трансляции завершается, полипептидная цепь покидает рибосому, транспортируется к специфическим органеллам клетки, где модифицируется в функционально активный белок.
IV.5. ГЕНЕТИЧЕСКИЙ КОД
Дж.Уотсон и Ф. Крик определили, что генетическая информация заключена в последовательности нуклеиновых оснований структуры ДНК. После того как было установлено, что синтез белка происходит в цитоплазме, стало очевидным, что должен существовать точный механизм переноса информации от ДНК, находящейся в ядре, к белку. Зависимость между последовательностью оснований ДНК ипоследовательностью аминокислотных остатков в белке получила название генетического кода. Если бы каждый отдельный из нуклеотидов кодировал одну аминокислоту, то белок мог включать только четыре аминокислоты. Если каждая аминокислота кодировалась бы только двумя основаниями, то это позволило бы закодировать 16 аминокислот. Однако лишь код, состоящий из трех оснований, может обеспечить включение в белок всех известных аминокислот. Подобный код содержит 64 различных сочетания из трех нуклеотидов. Доказательства триплетности генетического кода были впервые получены Ф. Криком в экспериментах с фагом Т4. В конце пятидесятых годов два ученых, М. Ниренберг и Г. Мат-теи, искусственно получили (синтезировали) РНК, состоящую из многократно повторяющегося урацила (поли-У). Это соединение (полиуридиловая кислота) было использовано в качестве мРНК, В каждую из 20 пробирок (по числу известных аминокислот) был внесен бесклеточный экстракт Е. coli, содержащий все необходимые компоненты для синтеза белка (рибосомы, тРНК, АТФ, другие ферменты), и одна из аминокислот. Затем в каждую пробирку добавляли поли-У. Анализ содержимого пробирок показал, что полипептид образовался только в той пробирке, которая содержала аминокислоту фенилаланин. Таким образом, было доказано, что триплет, или кодон УУУ, входящий в мРНК, определяет включение в полипептид аминокислоты фенилаланин. Аналогичные опыты показали, что триплет ЦЦЦкодирует аминокислоту пролин, а типлет ААА — лизин. Это открытие явилось первым шагом к расшифровке генетического кода. К 1965 г. генетический код был полностью расшифрован (рис. IV. 13). Из 64 кодонов три кодона УАГ, УАА, УГА не кодируют аминокислот и поэтому были названы бессмысленными или нонсенс-кодонами. Позднее оказалось, что они являются терминирующими кодонами. Назовем основные свойства генетического кода.
Рис. IV. 13. Генетический код
1. Генетический код триплетен. Триплет (кодон) — это последовательность трех нуклеотидов, кодирующая одну аминокислоту. 2. Генетический код является вырожденным: данная аминокислота может кодироваться более чем одним кодоном. Вырожденность кода обусловлена тем, что у кодонов, определяющих одну и ту же аминокислоту, первые два основания фиксированы, а третье положение может занимать другое основание. Исключение составляют метионин и триптофан, которые кодируются только одним триплетом. Триплет, соответствующий метионину (АУГ), инициирует считывание и не кодирует аминокислоту, если стоит в начале цепи ДНК. 3. Генетический код однозначен. Каждому кодону соответствует только одна аминокислота, т.е. триплет шифрует только одну аминокислоту. 4. Генетический код не перекрываем — процесс считывания генетического кода не допускает возможности перекрывания кодонов. 5. Генетический код универсален: одни и те же триплеты кодируют одни и те же аминокислоты у всех живых существ на Земле независимо от уровня их организации.
IV.6. ГЕНЫ И ИХ ОРГАНИЗАЦИЯ
Элементарной единицей наследственности является ген. Термин «ген» был предложен в 1909 г. В. Иогансеном для обозначения материальной единицы наследственности, выделенной Г.Менделем. После работ американских генетиков Дж. Бидла и Э. Тейтума геном стали называть участок молекулы ДНК, кодирующий синтез одного белка. Согласно современным представлениям, ген рассматривается как участок молекулы ДНК, характеризующийся специфической последовательностью нуклеотидов, определяющих аминокислотную последовательность полипептидной цепи какого-либо белка или нуклеотидную последовательность функционирующей молекулы РНК (тРНК, рРНК).
IV.6.1. Классификация генов
В зависимости от локализации генов в структурах клетки различают ядерные и митохондриальные гены (рис. IV. 14).
Рис. IV. 14. Классификация генов
По своему функциональному назначению гены могут быть разделены на две группы. Первая группа представлена генами, кодирующими белки; вторая группа — генами, контролирующими синтез РНК. Среди генов, кодирующих белки, различают: — гены «домашнего хозяйства», продукты которых необходимы для обеспечения функции любого типа клеток; — гены терминальной дифференцировки, т.е. гены, обеспечивающие специализированные функции клеток; — гены траскрипционных факторов, контролирующие особые ядерные белки, способные соединяться с регуляторными областями многих структурных генов, вызывая либо активацию, либо подавление транскрипции. РНК-кодирующие гены определяют синтез различных видов РНК, необходимых для синтеза рибосом, обеспечения процессов трансляции, сплайсинга, а также для синтеза молекул РНК, влияющих на функционирование других генов (регуляторное действие). Гены человека, как правило, представляют собой функционально прерывистую последовательность нуклеотидов (рис. IV. 15). Относительно короткие кодирующие последовательности оснований чередуются в них с длинными некодирующими последовательностями. Последовательности гена, представленные в молекуле зрелой иРНК, получили название экзонов. Именно экзоны являются кодирующими участками гена, контролирующими аминокислотную последовательность белков. Экзоны разделены некодирующими участками — нитронами, которые вырезаются (сплайсинг) в процессе созревания иРНК и не участвуют в процессе трансляции. В настоящее время в понятие «ген» включаются не только транскрибируемые области (экзоны и интроны), но и фланкирующие ген последовательности.
Рис. IV. 15. Схема организации гена
Фланкирующие области гена, как правило, высоко консервативны, т.е. характеризуются постоянством нуклеотидной последовательности, наблюдаемым даже при сравнении представителей различных видов. Фланкирующие области гена содержат последовательности, необходимые для его правильной работы: например, промоторная область в начале 5'-области или хвостовая нетранслируемая область поли-А, расположенная на З'-конце гена. Так, ТАТА — бокс (последовательность чередования тимина и аденина) обеспечивает правильную ориентацию РНК-полимеразы, что необходимо для транскрипции РНК. Размеры генов человека могут колебаться от нескольких десятков пар нуклеотидов (п. н.) до многих тысяч и даже миллионов п. п. Так, самый маленький из известных генов содержит всего 21 п. н., а один из самых крупных генов — ген дистрофина — имеет размер более 2, 6 млн п.н.
IV.7. ГЕНЫ И БЕЛКИ
После многолетних исследований серповидноклеточной анемии (малокровия) стало очевидным, что мутация определенного гена приводит к изменениям химической структуры молекулы гемоглобина. Основной тип гемоглобина взрослого человека состоит из четырех полипептидных (белковых) цепей: двух идентичных а-цепей и двух идентичных β -цепей (α 2β 2). В 1957 г. В. Ингрем обнаружил, что нормальный и серповидноклеточный гемоглобины содержат одинаковые а-цепи, но различные β -цепи. Различия между нормальной и мутантной (З-цепью заключались в том, что в серповидноклеточном гемоглобине остаток глутаминовой кислоты в 6-м положении заменен на остаток валина. Таким образом, различия между нормальным и мутантным вариантом, вызывающим заболевание, явились следствием молекулярного события — замены единственной аминокислоты в соответствующем белке. Выяснение природы серповидноклеточной анемии дало название целому классу заболеваний человека — «молекулярные болезни».
IV.7.1. Белки
Белки играют важнейшую роль в жизнедеятельности организмов. Каждый белок обладает своей уникальной функцией, которая определяется присущей только ему структурой и химическими свойствами. Выделяют следующие функции белков: ферментативную, структурную, двигательную, транспортную, питательную, защитную и регуляторную.
IV.7.2. Схема синтеза белка
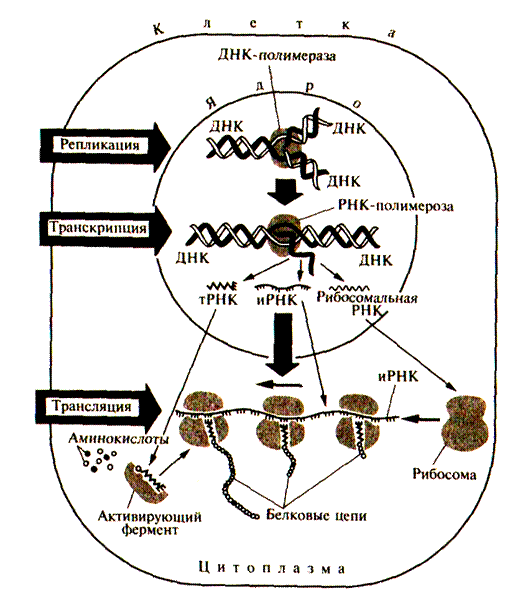
Процессы реализации информации, закодированной в специфической последовательности нуклеотидов конкретного гена, можно представлять обобщенно в так называемой центральной догме молекулярной биологин. Согласно ей одиночные цепи ДН К служат матрицами при синтезе комплементарных молекул (репликация). Далее смысловая цепь ДНК конкретного гена служит матрицей для синтеза точного транскрипта (пре-мРНК) соответствующего гена (транскрипция). Затем следует процесс созревания матричной РНК (процессинг), при котором происходит модификация молекулы и сплайсинг. Описанные события происходят в ядре клетки (рис. IV. 16). Зрелая и РНК выходит в цитоплазму, где на рибосомах осуществляется процесс перевода информации, записанной в последовательности иРНК, в аминокислотную последовательность соответствующего белка (трансляция).
Рис. IV.16. Схема репликации, транскрипции и трансляции генетической информации
Популярное:
|
Последнее изменение этой страницы: 2016-03-25; Просмотров: 1293; Нарушение авторского права страницы