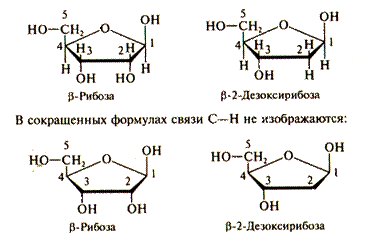|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
IV.2. ХИМИЧЕСКОЕ СТРОЕНИЕ НУКЛЕИНОВЫХ КИСЛОТ
Нуклеиновая кислота представляет собой гигантскую полимерную молекулу, построенную из многочисленных повторяющихся мономерных звеньев, называемых нуклеотидами.
Рис. IV.2. Строение и составные части нуклеотида
Нуклеотид состоит из азотистого основания, сахара и остатка фосфорной кислоты (рис. IV.2). Азотистые основания представляют собой производные одного из двух классов соединений — пуринового или пиримидинового ряда (рис. IV.3). В нуклеиновых кислотах присутствуют два производных пурина — аденин (обозначаемый А) и гуанин (Г) и три производных пиримидина — цитозин (Ц), тимин (Т) и урацил (У). В состав ДНК входят аденин, гуанин, цитозин и тимин. В структуре РНК тимин заменен другим пиримидином — урацилом. Сахар, входящий в состав нуклеотида, содержит пять углеродных атомов, т.е. представляет собой пентозу, которая может присутствовать в одной из двух форм: рибозы и дезоксирибозы (рис. IV. 4). Различие между ними состоит в том, что атом водорода при втором атоме углерода дезоксирибозы в рибозе замещен гидроксильной группой (—ОН). Сахар присоединяется к одному из оснований с помощью гликозидной связи, соединяющей 1-й углеродный атом пентозы с 1-м атомом азота производных пиримидина или с 9-м атомом азота производных пурина. В зависимости от формы пентозы различают два типа нуклеиновых кислот — дезоксирибонуклеиновую кислоту (ДНК) и рибонуклеиновую кислоту (РНК). Рис. IV.4. Строение рибозы и дезоксирибозы
Рис. IV.5. Схема формирования остова нуклеиновой кислоты
Нуклеиновые кислоты являются кислотами потому, что в их молекулу входит остаток фосфорной кислоты (—НРО3).В результате взаимодействия молекулы фосфорной кислоты с 3'-гидроксильной группой предшествующего нуклеотида и 5'-гидроксильной группой последующего образуется основная цепь нуклеиновой кислоты ( фосфат—сахар—фосфат—сахар и т.д.), к которой присоединяются разные азотистые основания (рис. IV.5). Порядок следования оснований вдоль цепи носит название первичной структуры нуклеиновой кислоты. Последовательность оснований определяется в направлении от 5'- к З'-углеродному атому пентозы.
IV.2.1. Строение ДНК
Пространственная конфигурации молекул ДНК была установлена в 1953 г. Это открытие было удостоено высшей научной награды — Нобелевской премии. Согласно модели, предложенной Дж. Уотсоном и Ф. Криком, молекула ДНК представляет собой две параллельные неразветвленные полинуклеотидные цепи, закрученные вокруг обшей оси в двойную правостороннюю спираль. Диаметр двойной спирали равен 20 им с шагом около 3, 4 нм, на каждыйвиток приходится 10 пар нуклеотидов, поскольку расстояние между соседними парами составляет 0, 34 нм (рис. IV.6). Направление цепей взаимно противоположно. На 5'-конце цепи ДНК всегда расположен свободный фосфат у 5'-атома углерода; на противоположном 31 конце — свободная ОН-группа у З'-атома углерода. Сахарно-фосфатный остов располагается по периферии спирали, а азотистые основания находятся внутри ее. Между основаниями образуются специфические водородные связи (рис.IV.7). В 50-е годы было установлено, что в ДНК, выделенной из клеток различных организмов, молярная сумма пуриновых оснований (аденин + гуанин) всегда равна сумме пиримидиновых (цитозин + тимин). Содержание аденина равно содержанию тимина, а содержание гуанина — количеству цитозина (правило Э.Чаргаффа).
Рис. IV.6. Схема строения молекулы ДНК
Рис. 1V.7. Комплементарность цепей в молекуле ДНК
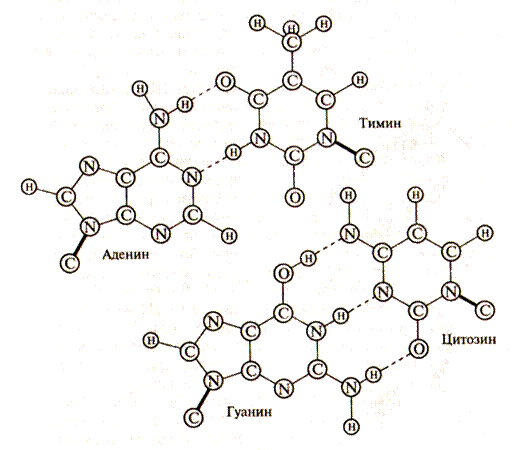
Рис. 1V.8. Спаривание аденина с тимином и гуанина с цитозином
Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи в результате избирательного спаривания оснований. Аденин всегда образует водородные связи с тимином (А-Т), а гуанин с цитозином (Г-Ц). Правило образования двунитевой структуры ДНК (А-Т, Г-Ц) называется правилом комплементарности, (лат. complementum — дополнение). Образование водородных связей между комплементарными парами обусловлено их пространственным соответствием (рис. IV.8). Это приводит к тому, что нуклеотидные последовательности двух антипараллельных цепей ДНК всегда строго комплементарны друг к другу, а порядок чередования нуклеотидов в обеих цепях ДНК оказывается взаимообусловленным. Именно комплеменарностью определяется точное воспроизведение последовательности оснований при копировании (репликации) молекул ДНК. Водородные связи между парами комплементарных нуклеотидов (две для пары А-Т и три для пары Г-Ц) относительно непрочные. Поэтому комплементарные нити молекулы ДНК могут разделяться и соединяться вновь при изменении некоторых условий (например, изменении температуры или концентрации солей). Разделение двух цепочечной ДНК называется денатурацией, а обратный процесс — образование двухцепочечной структуры ДНК — гибридизацией. Очевидно, что в реакцию гибридизации могут вступать только комплементарные друг другу нуклеотидные последовательности одноцепочечных молекул. Цепь, содержащая информацию о строении белка (в направлении 5'— 3'), называется смысловой цепью, а комплементарная — антисмысловой. Антисмысловая цепь имеет большое значение при стабилизации структуры двойной спирали ДНК и участвует в процессах репликации и репарации (восстановления) поврежденных участков ДНК. Молекулы ДНК являются гигантскими полимерами. Единицами измерения длины молекулы приняты: пары нуклеотидов (п.н.), тысячи пар нуклеотидов — килобазы (кб), миллионы пар оснований — мегабазы (мб). У человека гаплоидный набор содержит 3, 2-109 пар нуклеотидов, или 3, 2 миллиарда пар оснований. Почти вся ДНК клетки содержится в ядре в виде 46 плотно упакованных, суперскрученных за счет взаимодействий с ядерными белками, структурах — хромосомах. Сравнительно небольшая часть ДНК (около 5 %) локализована в митохондриях.
Популярное:
|
Последнее изменение этой страницы: 2016-03-25; Просмотров: 1776; Нарушение авторского права страницы