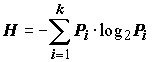|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Оценка качества экосистемы по индексам видового разнообразия
Видовое разнообразие слагается из двух компонентов: · видового богатства, или плотности видов, которое характеризуется общим числом имеющихся видов; · выравненности, основанной на относительном обилии или другом показателе значимости вида и положении его в структуре доминирования. Таким образом, один из главных компонентов биоразнообразия – видовое богатство или плотность видов – это просто общее число видов, которое в сравнительных целях иногда выражается как отношение числа видов к площади или числа видов к числу особей. Так, например, Р.Маргалеф предложил в качестве меры биоразнообразия индекс видового богатства Маргалефа:
d = (s – 1) / ln N, где s – число видов, N – число особей. Е.Ф. Менхиникк рассчитывал видовое богатство полевых насекомых по несколько другой формуле, используя в знаменателе функцию квадратного корня (индекс Менхиникка): dМ = (s – 1) / (N)1/2. Виды, входящие в состав биоценоза, очень сильно различаются по своей значимости. Традиционно принято выделение следующей иерархии видов: руководящие (или " доминантные" ) виды; за ними следует группа " субдоминантов"; остальные же виды считаются второстепенными, среди которых отмечают случайные или редкие. Значение отдельных видов должно определяться тем, какую роль играют они в функционировании экосистемы или в продукционном процессе. Но при исследованиях водных сообществ установить истинную функциональную роль видов нелегко, если об их значении судить только по обилию, т.е. численности и биомассе. При этом для анализа биоразнообразия и степени доминантности в разных ситуациях используют два традиционных подхода: · сравнения, основанные на формах кривых относительного обилия или доминирования – разнообразия: · сравнения, основанные на индексах разнообразия, представляющих собой отношения или другие математические выражения зависимости между числом видов и их значимостью. Е.Ф. Менхиникк рассчитывал видовое богатство полевых насекомых по несколько другой формуле, используя в знаменателе функцию квадратного корня (индекс Менхиникка): dМ = ( s – 1) / ( N )1/2. Индексы доминирования. Для природных биоценозов принято использовать индекс доминирования И. Балога: Di = Ni / Ns, где Ni – число особей i-го вида, Ns – общее число особей в биоценозе. К сожалению, этот идеальный по своей простоте индекс не отражает самого смысла доминирования, поскольку может принимать, например, значение 0.5 как при истинном доминировании, когда при нескольких сотнях видов один вид выражен половиной численности, так и в случае двух особей двух видов. Другая формула индекса доминирования (или доминантности) предложена А. Ковнацки на основе " коэффициента обилия" В.Ф. Палия (индекс доминирования Палия – Ковнацки):
Di = 100 × pi × Ni / Ns , (4.4)
где pi – встречаемость; pi = mi / Mi , mi – число проб, в которых был найден вид i, M – общее число проб, Ni – число особей i-го вида, Ns – общее число особей в биоценозе. Для характеристики видового комплекса предлагается выделять доминанты в пределах 10 < Di < 100, субдоминанты – в пределах 1 < Di < 10, субдоминанты первого порядка – в пределах 0.1 < Di < 1 и второстепенные члены – 0.01 < Di < 0.1. К. Шеннон определил энтропию опыта Н, как среднее значение неопределенности отдельных исходов: · для случая двух опытов H (a b ) = – (1/ r ) log(1/ r ) – (1/ l ) log(1/ l ) · или в общем случае произвольного опыта с k исходами, имеющими вероятности P1, P2, …, Pk
Энтропия (или неопределенность исхода) равна нулю, если вероятность одного из событий равна 1, и принимает максимальное значение в случае равновероятных исходов. Действительно, если известно, что в водоеме присутствует только один вид гидробионтов, то какая-либо неопределенность по его извлечению отсутствует (т.е. Н = 0). Неопределенность в предсказании результата отлова резко возрастает, если мы имеем в водоеме k видов с одинаковой численностью. Важным для биологии свойством энтропии является то, что значительным числом исходов, суммарная вероятность которых мала, при подсчете энтропии можно пренебречь. Энтропию H, как меру неопределенности, нельзя отождествлять с информациейI (как, например, напряженность электрического поля нельзя отождествлять с разностью потенциалов). Но количество информации об опыте b, содержащейся в опыте a, равно I ( a, b ) = H ( b ) – Ha ( b ) где Ha (b ) – условная энтропия опыта b после выполнения опыта a (т.е. снижение неопределенности b в результате выполнения a ). При расчете энтропии Н по Шеннону считается, что каждая проба – случайная выборка из сообщества, а соотношение видов в пробе отражает их реальное соотношение в природе. В качестве оценок вероятностей независимых событий рi для формулы (4.6) могут быть использованы следующие апостериорные отношения: · удельная численность i –го вида, как частное от деления его численности Ni на общую численность всех видов, взятых для анализа: pi = Ni / S Ni; · удельная биомасса i –го вида, как частное от деления его биомассы Bi на общую биомассу всех видов в пробе: pi = Bi / S Bi. Чуть позже Р. Маргалеф предложил другое выражение для индекса разнообразия:
где N! – факториальная величина всех исследуемых видов, ni! – факториал от числа особей каждого вида. Сопоставляя формулы, нетрудно увидеть, что формула Маргалефа – просто иная форма расчета энтропии по Шеннону. 13.12.3. Классификация водоемов и биоценозов
Классификация организмов по сапробности – это их классификация по сопротивляемости загрязнению (органической нагрузке, недостатку кислорода, присутствию соединений сероводорода), поскольку: сапробность (от греч. sapró s – гнилой) – «это комплекс физиологических свойств данного организма, обуславливающий его способность развиваться в воде с тем или иным содержанием органических веществ, с той или иной степенью загрязнения».. Для каждой зоны сапробности можно выделить тесно связанное с ней подмножество видов гидробионтов, которые считаются ее индикаторами. Именно это обстоятельство породило иллюзию того, что в основании сапробиологической классификации водоемов лежат именно " биологические" факторы, а не механизмы деструкции органического вещества. О.П. Оксиюк и В.Н. Жукинский в своих классификационных таблицах соотнесли две шкалы: сапробности и трофности. Если под сапробностью понимается интенсивность органического распада, то трофность означает интенсивность органического синтеза. В природе оба процесса – органический синтез и распад – существуют параллельно и состоят друг с другом в многократном взаимодействии, что позволяет говорить об аналогии ступеней сапробности и трофики: " олигосапробность – олиготрофия", " b-мезосапробность – мезотрофия", " a–мезасапробность – эвтрофия" и " полисапробность – гипертрофия". Эта аналогия привлекательна тем, что создает предпосылку к устранению одной из классификаций, как ненужного дублирующего звена. В худших конкурентных условиях находится система сапробности, как основанная на весьма " размытых" разделяющих факторах, когда как классификация по трофике жестко связана с концентрациями биогенных элементов. В то же время, ряд исследователей подчеркивает неполное совпадение форм трофики и сапробности, особенно в мезосапробных зонах и для непроточных водоемов. Система Кольквитца–Марссона была разработана применительно к условиям загрязнения вод средней Европы в начале века. В настоящее время характер и степень загрязнения водоемов изменились, в основном за счет интенсификации антропогенного воздействия. Это явилось причиной расширения " классической" классификации в двух основных направлениях: · появление новых зон " чище" олигосапробной и " грязнее" полисапробной; · выделение дополнительных зон на принципиально новой классификационной основе. Наиболее широкая ревизия " классической" системы была выполнена В. Сладечеком, который включил в классификацию абиотические зоны, а внутри полисапробной выделил три зоны – изосапробную (преобладание цилиат над флагеллятами), метасапробную (преобладание флагеллят над цилиатами) и гиперсапробную (отсутствие простейших при развитии бактерий и грибов). Наконец, была сделана методологически решительная попытка [Slá deč ek, I969М] сравнения некоторых бактериологических и химических показателей с отдельными ступенями сапробности и предложена общая " биологическая" схема качества вод. Все системы сапробности учитывают фактически только нетоксичные органические загрязнения, которые влияют на организмы в первую очередь через изменение кислородного режима. Для учета влияния токсических органических и неорганических соединений делаются попытки разработать шкалы токсобности и затем объединить их со шкалами сапробности в единую шкалу сапротоксобности, причем существуют противоположные мнения о возможности такого объединения. Популярное:
|
Последнее изменение этой страницы: 2016-03-25; Просмотров: 1531; Нарушение авторского права страницы