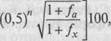|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Группы сцепления генов у кур (по И. А. Захарову, 1979)
Фенотип
Дефекты скелета ног и крыльев Перьев мало Голая шея Голубая скорлупа яиц Гороховидный гребень Гибель на поздней стадии инкубации Добавочные пальцы Сильно выраженная полидактилия Приступы столбняка Гребень розовидный Оперение серебристое Неполный альбинизм Веки полузакрытые Летальные расстройства нервной системы Раздвоение сосочка копчиковой железы Уменьшение крыльев в различной степени (дефекты развития) После определения локализации группы сцепления генов в конкретной паре гомологичных хромосом составляют карты хромосом. Для картирования генов в хромосоме необходимо установить сцепление не менее трех неаллельных генов. птицеводстве имеют сцепленные с полом признаки, которые контролируются генами, локализованными в Х-хромосоме. К таким генам относят: ген карликовости (dw), ген полосатого оперения (В), ген серебристого оперения (5), ген медленной оперяемости (Я), ген коричневых глаз (br) и др. В настоящее время у кур известно 1.5 мутантных генов, локализованных в Jif-хромосоме. Кроме того, установлено несколько, пока не локализованных в данной хромосоме генов, но сцепленных с полом: Ьа (плешивость), chz (хондродистрофия), he (гистоанти-ген), у (нервное возбуждение), ga (удушье) и др. В качестве примера использования закономерностей наследования сцепленных с полом признаков можно привести создание аутосексных (различаемых по полу в суточном возрасте) линий и кроссов кур (яичных — «Хайсекс коричневый», «Ломанн коричневый», «Росс белый», «Родонит»; мясных — «Кобб 100+», «Конкурент» и др.). Получение таких цыплят основано на разли-Первый случай сцепления аутосомных генов — розовидного гребня и коротконогое™ выявили в 1928 г. наши соотечественники А. С. Серебров-ский и С. Г. Петров. Важное практическое значение в
чиях в окраске и скорости оперения у суточных петушков и курочек. Особое значение селекционеры придают рецессивному гену dw. Он обусловливает уменьшение массы тела до 30 %. Выведены линии мини-кур мясного и яичного направлений продуктивности. Изменчивость признаков. Большинство хозяйственно полезных признаков у птицы легко изменяются под действием факторов внешней среды, и даже сходные генотипы в разных условиях могут иметь различные фенотипы. Это свидетельствует о том, что фенотип особи есть результат взаимодействия генетических и негенетических факторов. Изменчивость бывает генотипическая (наследственная) и модификационная (ненаследственная). Наследственную изменчивость подразделяют на онтогенетическую, комбинационную, мутационную и коррелятивную. Онтогенетическая изменчивость — это совокупность последовательных изменений признаков и свойств особи в процессе индивидуального развития (онтогенеза), когда каждый признак формируется самостоятельно, но в строгом соответствии с генетически детерминированным общим планом развития данной птицы. Комбинационная изменчивость возникает вследствие случайного сочетания генов отцовского и материнского организмов при слиянии половых клеток и образовании зиготы, а также в результате перегруппировки генов в хромосомах. При этом отмечено, что сами гены не изменяются, а изменяется их сочетание и характер взаимодействия. Таким образом, комбинационная изменчивость у птицы выражается сочетанием у потомства разных признаков того и другого родителя. Комбинационная изменчивость лежит в основе создания многих пород, линий, кроссов и типов сельскохозяйственной птицы. Достигают этого путем целенаправленного отбора и подбора птицы. Так, бройлеров основных кроссов мясных кур получают в результате скрещивания сочетающихся линий (комбинаций) пород кор-ниш и белый плимутрок. Мутационная изменчивость проявляется внезапно в результате изменений структуры генов и хромосом особи. Процесс образования мутаций называют мутагенезом, а факторы, вызывающие их? _ мутагенами. Мутации, возникающие под влиянием естественных факторов внешней среды, в результате физиологических и биохимических изменений в организме, называют спонтанными, а искусственно вызванные действием химических веществ, радиации, высоких температур и т. д. — индуцированными. Крайняя форма генной мутации — появление нового гена (нового признака), хромосомной мутации — образование нового вида хромосом, геномной мутации — образование нового генома на базе объединения и преобразования разных геномов двух или нескольких биологических видов.
Мутации бывают нейтральные, вредные и полезные (используемые при создании новых пород, линий и кроссов птицы). К нейтральным мутациям, не вызывающим летального исхода у птицы, относят следующие признаки: длинный хвост у петухов декоративных японских пород феникс и Иокогама; курчавость и шелковистость оперения у кур; курчавость оперения у гусей некоторых пород (например, севастопольской); красноголовость у японских перепелов и др. Вредные мутации вызываются, как правило, летальными и полулетальными генами в гомозиготном состоянии. В качестве примера можно привести аутосомную рецессивную мутацию, обусловливающую врожденный дефект органов равновесия у кур, индеек и японского перепела (вызывается геном 1о). Пораженный молодняк всех трех родственных видов может стоять несколько секунд, затем падает с повернутой набок головой, перестает есть, пить и погибает на 9-й день после вывода даже при самом заботливом уходе. Более важное значение для селекционеров имеют полезные мутации. К ним относят: карликовость яичных и мясных кур — носителей рецессивного, сцепленного с полом гена dw, на базе которых созданы и используются в промышленном птицеводстве мясные и яичные породы и линии мини-кур; сцепленная с полом доминантная мутация серебристости (ген S), на базе которой можно создавать аутосексных цыплят; ген с рецессивной белой окраски оперения, который специально вводят в генотип мясных кур для получения тушек (бройлеров) лучшего товарного вида и др. Модификационная изменчивость характеризуется изменением признаков организма (фенотипа) под влиянием факторов внешней среды. При модификации у одинаковых генотипов фенотипы изменяются. Примерами модификации признаков служат изменение яйценоскости птиц под влиянием длины светового дня и интенсивности освещения, изменение скорости роста и воспроизводительных качеств птиц в зависимости от кормления и т. д. Но изменчивость признаков организма может происходить лишь в пределах, ограниченных возможностями данного генотипа. В отличие от мутаций модификации чаще проявляются у многих животных, а не у отдельных особей, и при сходной наследственности имеют однонаправленный характер. При создании оптимальных условий кормления и содержания можно вести целенаправленную работу по формированию стад птицы с желательными признаками. Кроме вышеуказанных видов изменчивости важное значение для практического птицеводства имеет коррелятивная (соотносительная) изменчивость. Корреляция — зависимость между изменчивостью двух или нескольких признаков, то есть изменение одного признака ведет к изменению другого. В основе корреляции лежат генетические факторы, факторы внешней среды и физиологическое состояние организма. Различают следующие типы кор-
реляции: генетическую, вызванную плейотропным действием генов или их сцеплением; средовую, обусловленную факторами внешней среды; фенотипическую, возникающую на основе взаимодействия генотипа и среды. Корреляционная связь может быть положительной (например, между живой массой и массой яйца у кур), отрицательной (между высокой яйценоскостью и размером яиц) и нейтральной (между живой массой курочек и их половой зрелостью). Если коэффициент корреляции 0, 3 и ниже, то связь считают малой; 0, 31—0, 59 — средней; 0, 6 и выше—значительной. Зная уровень корреляции между признаками, ее направление и величину, можно проводить эффективнее отбор по одному или нескольким желательным признакам, прогнозировать изменение одних признаков при отборе птицы по другим. Корреляция может быть изменена в результате направленной селекции в стаде. Для характеристики изменчивости признака особей данной совокупности применяют среднее квадратическое отклонение (а), вариансу (а2), коэффициент изменчивости (Cv), нормированное отклонение (t), вариационный размах (хмакс — хмин). Инбридинг и инбредная депрессия. Инбридинг — это система спаривания птиц, состоящих между собой в более тесных родственных отношениях, чем это в среднем встречается в популяции (например, в породе, линии, стаде). В зависимости от степени родства спариваемых особей различают следующие типы инбридинга: кровосмешение (очень тесный инбридинг), при котором проводят спаривание по схеме мать х сын (такое спаривание по методу А. Шапоружа можно записать как I—II), дочь х отец (II—I), сестра х брат (II—II); близкий инбридинг I—III (бабка хвнук), III—I (внучках дед), II—II (полусибсы, двоюродные брат х сестра), II—III (тетя х племянник), III—II (племянницахдядя) и т.д.; умеренный инбридинг — общий предок встречается в III—IV, IV—HI, IV-—IV поколениях; отдаленный инбридинг — общий предок встречается в IV—V, V-IV, V-V, V-VI поколениях. При всех типах инбридинга происходят повышение гомозигот-ности потомства и генетическое расщепление, в результате которого инбредные группы получают различные наборы генов. Генетические последствия инбридинга заключаются в том, что в популяции гомозиготными становится большее число пар генов независимо от типа генного взаимодействия. Именно этим генетическим явлением обусловлены все фенотипические последствия инбридинга. Гомозиготность вызывает перекомбинацию генов, что ведет к изменению взаимодействия между генами различных локусов. Происходят так называемые эпистатические эффекты,
Известно, что сильнее всего инбридинг действует на те признаки, которые связаны с воспроизводительными качествами птицы и ее жизнеспособностью. Низкие плодовитость и жизнеспособность, передающиеся из поколения в поколение, обусловлены в первую очередь сужением благоприятного взаимодействия генов внутри локусов и между ними, что зачастую приводит к летальному исходу. Необходимо отметить, что все эти признаки слабо наследуются. Явление, при котором в результате инбридинга снижаются продуктивность и жизнеспособность птицы (резистентность организма), называют ынбредной депрессией. Это связано с выщеплени-ем в гомозиготном состоянии отрицательно действующих генов, в том числе летальных, а также с нарушением сбалансированности полигенной системы. Сила проявления инбредной депрессии зависит от следующих факторов: видовой, породной и линейной принадлежности птицы (например, мясные породы значительно менее устойчивы, чем яичные); индивидуальных особенностей инбридируемых птиц, степени их исходной гетерозиготности, конституциональной крепости и т.д.; пола (самцы более подвержены инбредной депрессии, чем самки); возраста (при спаривании особей в раннем возрасте и старости инбредная депрессия проявляется сильнее); природы признака (чаще всего признаки, отличающиеся высокой степенью наследуемости — живая масса, масса яиц — менее подвержены инбредной депрессии, чем низко- и средненаследуе-мые — выводимость, выживаемость, яйценоскость); скорость падения гетерозиготности и числа инбридированных поколений (чем теснее инбридинг, тем, естественно, быстрее с каждым поколением снижается гетерозиготность и тем сильнее проявляется инбредная депрессия); условий среды (оптимальные для выживания условия среды способствуют ослаблению инбредной депрессии, повышению продуктивности птицы, то есть лучшей реализации ее генетических возможностей). Таким образом, переход отрицательно действующих (нежелательных) генов в гомозиготное состояние способствует выявлению и удалению их из стада. Особенно эффективно в этом направлении действие инбридинга в отношении очищения стада от летальных и полулетальных генов. Поэтому в птицеводстве инбридинг часто выполняет «санитарную роль». По обобщенным данным, применение тесного инбридинга в ряде поколений приводит к снижению яйценоскости кур на 8—17 % (в среднем 11), выводимости яиц на 12—17 (в среднем 14, 5), вывода цыплят на 4—
10 (в среднем 7, 5), жизнеспособности молодняка и взрослых кур на 7—12 (в среднем 9, 5) и скорости роста мясных цыплят на 3—6 % (в среднем 4, 4). В то же время на массу яйца инбридинг практически не оказывает отрицательного воздействия. Так, при повышении коэффициента инбридинга на 1 % продуктивность кур яичных линий снижается на одно яйцо выводимость яиц на 0, 53 и сохранность птицы на 0, 55 % (Киселев Л. Ю., Фатеев В. Н., 1983). Для расчета коэффициента инбридинга существует несколько методов. В птицеводстве наиболее распространен (помимо метода А. Шапоружа) метод С. Райта — Д. Кисловского, при котором используют следующую формулу: где Fy■ — коэффициент инбридинга, %; X — знак суммирования коэффициентов инбридинга разных общих предков; п — число рядов предков от общего предка по материнской стороне родословной; г\\ — число рядов предков от общего предка по отцовской стороне родословной; fa — коэффициент инбридинга для общего предка, выраженный в долях единицы. При использовании данной формулы отсчет рядов предков начинают с родительского ряда. Если среди общих предков не встречаются инбридированные особи, то коэффициент инбридинга вычисляют по упрощенной формуле: Пример расчета коэффициента инбридинга петуха № 1401
Общие предки петуха № 1401 —курица № 1215 и петух №2503. Используя формулу С. Райта — Д. Кисловского, получим, что коэффициент инбридинга у петуха № 1401 составляет 15, 625 %. Fx = [(0, 5)2+2-] + (О^)3^-1] 100 = [(0, 5)3 + (0, 5)5] 100 * (0, 125 + 0, 03125)100 - = 15, 625%. По А. Шапоружу родство по предку № 2503 записывают как II—II, а по предку № 1215 — III—III. Таким образом, петух № 1401 был выведен с использованием близкого инбридинга. При коэффициенте 25 % и более инбридинг считается очень тесным (кровосмешение); от 12, 5 до 25 —близким; от 1, 55 до 12, 5 — умеренным; от 0, 20 до 1, 55 % — отдаленным.
Птица, у которой один или несколько общих предков, родственна между собой и, естественно, имеет генетическое сходство, то есть сходную наследственность. Причем она тем больше, чем ближе в рядах родословных расположен общий предок. Если же общий предок инбридирован, то генетическое сходство будет еще выше. Генетическое сходство потомков с выдающимся предком, на которого ведется селекция, расчитывют по следующей формуле:
где Rxcl — коэффициент генетического сходства между животным (х) и его предком (а); я —ряды в родословной животного, в которых встречается предок; /, — коэффициент инбридинга предка; /х — коэффициент инбридинга самого животного. Вычисление коэффициента генетического сходства аналогично вычислению коэффициента инбридинга. На возрастание гомозиготности (коэффициента инбридинга) значительное влияние оказывают продолжительность и тип родственного спаривания (табл. 23).
23. Возрастание гомозиготности в поколениях в зависимости от типа спаривания, %
Возвратное спаривание на инбридированных отца или мать о инбридинг увеличивает число пар генов, которые становятся
Ввиду того чт гомозиготными независимо от их фенотипической экспрессии и числа, многие ученые в области генетики и селекции сделали определенные выводы относительно важности генетических последствий инбридинга. Особого внимания заслуживают высказывания Д. Лэсли (1982) о генетических последствиях гомозиготности. 1. Инбридинг, или возрастание гомозиготности, не увеличива 2. Инбридинг не раскрывает доминантные гены, так как гомо
готными, чем гетерозиготными. Зная, что доминантные гены обычно обладают благоприятным эффектом, а рецессивные — неблагоприятным, выбраковка нежелательных животных должна в конечном итоге привести к увеличению благоприятных (доминантных) генов в популяции. Поэтому при использовании близкородственного спаривания необходимо проводить тщательный отбор животных и жесткую выбраковку менее желательных особей в стаде. 3. Инбридинг способствует закреплению признаков в инбред- 4. Родственное спаривание может способствовать увеличению 5. Возрастание степени гомозиготности сопровождается сниже Однако увеличение гомозиготности, которое теоретически можно ожидать при разных типах инбридинга, на практике оказывается часто невыполнимым вследствие наступления инбред-ной депрессии. Инбридинг в разных формах используют в селекции, так как с его помощью можно выявить имеющиеся в популяции (стаде) ценные комбинации генов и закрепить их в потомстве. Инбридинг способствует закреплению высоких качеств выдающихся производителей в потомстве, то есть преобразованию полезных качеств отдельных особей в групповые, свойственные линии. Линии, созданные на основе тесного инбридинга в течение ряда поколений, называют инбредными. Они характеризуются высокой степенью гомозиготности, большей генетической однородностью индивидов, чем особи гетерогенной популяции. Кроме того, птицу инбредной линии отличает высокая однородность особей по морфологическим и физиолого-биохимическим признакам. Для получения инбредных линий, как правило, применяют тесный инбридинг по типу брат х сестра в течение 4—5 поколений, при этом жизнеспособность и продуктивность птицы резко падают. Поэтому для дальнейшего воспроизводства отбирают лишь небольшое поголовье птицы, выдержавшей инбредную депрессию, что дает возможность сохранить уровень продуктивности, свойственный птице исходных линий. Испытания создаваемых инбредных линий на сочетаемость целесообразно начинать при коэффициенте инбридинга 12—15 %..
При последующих скрещиваниях сочетающихся инбредных линий между собой (межлинейные кроссы) получают гетерозиготное потомство в I поколении по одному или нескольким экономически важным признакам. Однако такой путь создания кроссов для промышленного птицеводства требует значительных средств и времени, так как очень велик отход инбредных линий, а количество сочетающихся линий среди оставшихся мало. В связи с этим для получения гибридной птицы в промышленном птицеводстве используют специализированные сочетающиеся линии отцовских и материнских форм, о которых подробно сказано ниже. Без существенного отрицательного влияния на продуктивность и жизнеспособность птицы коэффициент инбридинга можно довести в линиях яичных кур до 35—40 %, в линиях мясных кур до 12—15, в линиях индеек до 36—39%. Значительное повышение степени инбридинга приводит к инбредной депрессии с ослаблением конституции, снижением жизнеспособности, воспроизводительных и продуктивных качеств птицы, что существенно ограничивает возможности использования инбридинга в практической селекции. Поэтому при работе с линиями птицы в племенных хозяйствах с целью недопущения вредных последствий инбридинга необходимо избегать близкородственного спаривания, если это не предусматривается специальной селекционной программой. Важными мерами борьбы с вредными последствиями инбридинга служат строгий отбор для племенных целей особей с крепкой конституцией, использование метода циклической селекции, создание оптимальных условий кормления и содержания для самцов и самок. Гетерозис. Под гетерозисом понимают сложное биологическое явление, при котором птица, полученная от скрещивания при определенном подборе, превосходит лучшую из родительских форм по жизнеспособности, энергии роста, плодовитости, крепости конституции, устойчивости к заболеваниям и продуктивности. Многие ученые считают проявлением эффекта гетерозиса и те случаи, когда гибридное потомство, полученное в результате скрещивания, превышает по одному или нескольким хозяйственно полезным признакам средние показатели между родительскими формами, а также одну из родительских форм, не превосходящую лучшую из них. Гетерозис проявляется у потомства I поколения, затем он заметно затухает и исчезает в последующих поколениях, если не применять различные виды скрещивания, при которых учитываются новые комплексы генотипов. В основе гетерозиса лежат: наследование количественного характера различных признаков; важнейшие генетические и биологические процессы, происходящие в организме (различные формы взаимодействия генов между собой и генных продуктов с цитоплаз-
мой, между генами и средой и т. д.); более интенсивное протекание нуклеинового обмена; активность тканевых ферментов и повышение интенсивности окислительно-восстановительных процессов в организме; 5) улучшение функционирования пищеварительной системы и органов размножения, что в конечном итоге способствует повышению уровня хозяйственно полезных признаков птицы. С развитием генетики был выдвинут ряд гипотез, объясняющих эффект гетерозиса. 1. Гипотеза доминантных генов. Эффект гетерозиса согласно 2. Гипотеза сверхдоминирования. Суть ее состоит в том, что ге- 3. Гипотеза облигатной гетерозиготности. Разработана она
генами с двойным действием: полезным (доминантным) и вредным (рецессивным). Возникновение таких генов является следствием эволюционного процесса. В процессе эволюции выживают те особи, у которых полезные гены сохраняются в доминантном состоянии, а вредные — в рецессивном. 4. Гипотеза генетического баланса. Под генетическим балансом подразумевают сбалансированность всего генома (комплекса ДНК, заключенного в одном наборе хромосом), влияющего на развитие организма в целом. Согласно этой теории эффект гетерозиса обусловлен влиянием большого числа генов, которые в определенной степени сбалансированы в геноме, под воздействием естественного или искусственного отбора. При скрещивании особей с определенными геномами происходит образование новой комбинации генома у потомства, вызывающей гетерозис. В то же время при родственном спаривании особей новая комбинация генома у потомства сопровождается, как правило, снижением жизнеспособности и плодовитости. Таким образом, скрещивание и инбридинг нарушают генетический баланс и приводят к гетерозису либо к инбредной депрессии. На проявление гетерозиса влияют в основном те же факторы, что и при инбридинге, однако их действие различно. Если оптимальные условия среды способствуют преодолению инбредной депрессии, то аналогично они приводят к большей эффективности проявления гетерозиса. С другой стороны, природа признаков, их наследуемость влияют разнонаправленно на проявление инбридинга и гетерозиса. Низконаследуемые признаки наиболее сильно подвержены инбредной депрессии, однако по ним больше всего проявляется эффект гетерозиса. По высоконаследуемым признакам этот эффект практически не проявляется (табл. 24). Популярное:
|
Последнее изменение этой страницы: 2016-03-25; Просмотров: 2125; Нарушение авторского права страницы
 Продолжение
Продолжение Мраморный узор на пухе у цыплят
Мраморный узор на пухе у цыплят