|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ЗАКОНОМЕРНОСТИ ИНДИВИДУАЛЬНОГО РАЗВИТИЯ ОРГАНИЗМОВ
ОСНОВНЫЕ КОНЦЕПЦИИ В БИОЛОГИИ ИНДИВИДУАЛЬНОГО РАЗВИТИЯ
В предыдущих главах были рассмотрены генетические закономерности, определяющие формирование определенного фенотипа, изложено основное содержание стадий онтогенеза, последовательно и закономерно сменяющих друг друга. Все эти сведения не дают, однако, ответа на вопрос, почему и каким образом генотип реализуется в фенотип в виде тех или иных клеточных и системных процессов, в виде сложных пространственных и упорядоченных во времени онтогенетических преобразований. При сравнении зиготы и половозрелой особи, которые, по сути, являются двумя разными онтогенетическими стадиями существования одного и того же организма, обнаруживаются очевидные различия, касающиеся по крайней мере размеров и формы. Начиная с XVII в. ученые пытались познать и объяснить процессы, приводящие к этим количественным и качественным изменениям особи. Первоначально возникла гипотеза, согласно которой онтогенез рассматривали лишь как рост расположенных в определенном пространственном порядке предсуществующих структур и частей будущего организма. В рамках этой гипотезы, получившей название преформизма, каких-либо новообразований или преобразований структур в индивидуальном развитии не происходит. Логическое завершение идеи преформизма заключается в допущении абсурдной мысли о «заготовленности» в зиготе и даже в половых клетках прародителей структур организмов всех последующих поколений, как бы вложенных последовательно наподобие деревянных матрешек. Альтернативная концепция эпигенеза была сформулирована в середине XVIII в. Ф. К. Вольфом, впервые обнаружившим новообразование нервной трубки и кишечника в ходе эмбрионального развития. Индивидуальное развитие стали связывать целиком с качественными изменениями, полагая, что структуры и части организма возникают как новообразования из бесструктурной яйцеклетки. В XIX в. К. Бэр впервые описал яйцо млекопитающих и человека, а также зародышевые листки и обнаружил сходство плана строения зародышей различных классов позвоночных — рыб, амфибий, рептилий, птиц, млекопитающих. Он же обратил внимание на преемственность в этапах развития — от более простого к более сложному. Бэр рассматривал онтогенез не как предобразование, не как новообразование структур, а как их преобразование, что вполне согласуется с современными представлениями. Выяснение конкретных клеточных и системных механизмов таких преобразований составляет основную проблему современной биологии развития. Увеличение массы тела особи, т.е. ее рост, и появление новых структур в ходе ее развития, называемое морфогенезом, нуждаются в объяснении. Рост и морфогенез подчиняются законам, которые обусловливают приуроченность конкретных процессов онтогенеза к определенному месту зародыша и периоду эмбриогенеза. Отдельные стадии индивидуального развития отличаются также определенной скоростью протекания с характерным качественным и количественным результатом. Биология развития изучает способы генетического контроля индивидуального развития и особенности реализации генетической программы в фенотип в зависимости от условий. Под условиями понимают различные внутриуровневые и межуровневые процессы и взаимодействия: внутриклеточные, межклеточные, тканевые, внутриорганные, организменные, популяционные, экологические. Можно сказать, что усилия исследователей в области биологии развития концентрируются вокруг стержневой проблемы генетической предопределенности и лабильности онтогенетических процессов, что в известном смысле на ином уровне познания возвращает нас к идеям неопреформизма и эпигенеза. Не менее важными являются исследования конкретных онтогенетических механизмов роста и морфогенеэа. К ним относятся следующие процессы: пролиферация, или размножение клеток, миграция, или перемещение клеток, сортировка клеток, их запрограммированная гибель, дифференцировка клеток, контактные взаимодействия клеток (индукция и компетенция), дистантные взаимодействия клеток, тканей и органов (гуморальные и нервные механизмы интеграции). Все эти процессы носят избирательный характер, т.е. протекают в определенных пространственно-временных рамках с определенной интенсивностью, подчиняясь принципу целостности развивающегося организма. Биология развития стремится выяснить степень и конкретные пути контроля со стороны генома и одновременно уровень автономности различных процессов в ходе онтогенеза.
МЕХАНИЗМЫ ОНТОГЕНЕЗА
Деление клеток
Деление клеток играет большую роль в процессах онтогенеза. Во-первых, благодаря делению из зиготы, которая соответствует одноклеточной стадии развития, возникает многоклеточный организм. Во-вторых, пролиферация клеток, происходящая после стадии дробления, обеспечивает рост организма. В-третьих, избирательному размножению клеток принадлежит заметная роль в обеспечении морфогенетических процессов. В постнатальном периоде индивидуального развития благодаря клеточному делению осуществляется обновление многих тканей в процессе жизнедеятельности организма, а также восстановление утраченных органов, заживление ран. Зигота, бластомеры и все соматические клетки организма, за исключением половых клеток, в периоде созревания гаметогенеза делятся митозом. Клеточное деление как таковое является одной из фаз клеточного цикла. От продолжительности интерфазы (G1 + S + G2-периоды) зависит частота последовательных делений в ряду клеточных поколений. В свою очередь интерфаза имеет разную продолжительность в зависимости от стадии развития зародыша, локализации и функции клеток. Так, в периоде дробления эмбриогенеза клетки делятся быстрее, чем в другие, более поздние периоды. Во время гаструляции и органогенеза клетки делятся избирательно в определенных областях зародыша. Замечено, что там, где скорость клеточного деления высокая, происходят и качественные изменения в структуре эмбриональной закладки, т.е. органогенетические процессы сопровождаются активным размножением клеток. Показано, что растяжение клеток при их движении стимулирует клеточное деление. В сформировавшемся организме некоторые клетки, например нейроны, вообще не делятся, в то время как в кроветворной и эпителиальной тканях продолжается активное размножение клеток. Клетки некоторых органов взрослого организма в обычных условиях почти не делятся (печень, почка), но при наличии стимула в виде воздействия гормональных или внутритканевых факторов, часть из них может вступить в деление. При изучении расположения делящихся клеток в тканях обнаружено, что они группируются гнездами. Само по себе деление клеток не придает эмбриональному зачатку определенной формы, и нередко эти клетки располагаются беспорядочно, но в результате последующего их перераспределения и миграции зачаток приобретает форму. Так, например, в зачатке головного мозга деление клеток сосредоточено исключительно в том слое стенки, который прилежит к полости невроцеля. Затем клетки передвигаются из зоны размножения к наружной стороне пласта и образуют ряд выпячиваний, так называемых мозговых пузырей. Таким образом, клеточное деление в эмбриогенезе носит избирательный и закономерный характер. Об этом же свидетельствует открытая в 60-х годах суточная периодичность количества делящихся клеток в обновляющихся тканях. В настоящее время известен ряд веществ, которые побуждают клетки к делению, например фитогемагглютинин, некоторые гормоны, а также комплекс веществ, выделяющихся при повреждении тканей. Открыты также и тканеспецифичные ингибиторы клеточного деления — кейлоны. Их действие заключается в подавлении или замедлении скорости деления клеток в тех тканях, которые их вырабатывают. Например, эпидермальные кейлоны действуют только на эпидермис. Будучи тканеспецифичными, кейлоны лишены видовой специфичности. Так, эпидермальный кейлон трески действует и на эпидермис млекопитающего. За последние годы установлено, что многие структуры зародыша образуются клетками, происходящими от небольшого числа или даже одной клетки. Совокупность клеток, являющихся потомками одной родоначальной клетки, называют клоном. Показано, например, что большие по объему участки центральной нервной системы формируются из определенных клеток раннего зародыша. Пока не ясно, в какой именно срок происходит отбор родоначальных клеток, каков механизм этого отбора. Важным следствием такой селекции является то, что многим клеткам раннего зародыша не суждено участвовать в дальнейшем развитии. В опытах на мышах показано, что организм развивается всего из трех клеток внутренней клеточной массы на стадии, когда бластоциста состоит из 64 клеток, а сама внутренняя клеточная масса содержит примерно 15 клеток. Клональные клетки могут быть причиной мозаицизма, когда большие группы клеток отличаются по набору хромосом или аллельному составу. По-видимому, количество циклов клеточных делений в ходе онтогенеза генетически предопределено. Вместе с тем известна мутация, изменяющая размеры организма за счет одного дополнительного клеточного деления. Это мутация gt (giant), описанная у Drosophila melanogaster. Она наследуется по рецессивному сцепленному с полом типу. У мутантов gt развитие протекает нормально на протяжении всего эмбрионального периода. Однако в тот момент, когда нормальные особи окукливаются и начинают метаморфоз, особи gt продолжают оставаться в личиночном состоянии еще дополнительно 2—5 сут. За это время у них происходит одно, а может быть, и два дополнительных деления в имагинальных дисках, от количества клеток которых зависит размер будущей взрослой особи. Затем мутанты образуют куколку вдвое крупнее обычной. После метаморфоза несколько удлиненной по времени стадии куколки на свет появляется морфологически нормальная взрослая особь удвоенного размера. У мышей описан ряд мутаций, обусловливающих снижение пролиферативной активности и следующие за этим фенотипические эффекты. К ним относят, например, мутацию or (ocular retardation), затрагивающую сетчатку глаза начиная с 10-х суток эмбрионального развития и приводящую к микрофтальмии (уменьшению размеров глазных яблок), и мутацию tgia, затрагивающую центральную нервную систему с 5—6-х суток после рождения и приводящую к отставанию роста и атрофии некоторых внутренних органов. Таким образом, деление клеток является чрезвычайно важным процессом в онтогенетическом развитии. Оно протекает с разной интенсивностью в разное время и в разных местах, носит клональный характер и подвержено генетическому контролю. Все это характеризует клеточное деление как сложнейшую функцию целостного организма, подчиняющегося регулирующим влияниям на различных уровнях: генетическом, тканевом, онтогенетическом. Миграция клеток
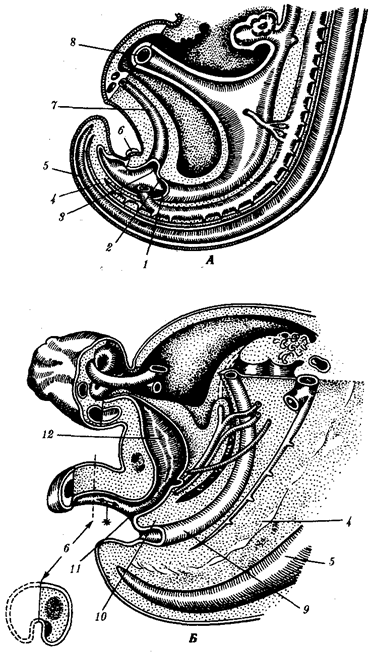
Миграции клеток, или клеточные перемещения, наряду с другими клеточными процессами имеют очень большое значение, начиная с процесса гаструляции и далее, в процессах морфогенеза. Клетки мезенхимного типа мигрируют одиночно и группами, а клетки эпителиев обычно согласованно, пластом. Мезенхима — это скопление веретеновидных или звездчатых клеток, погруженных в межклеточный матрикс. Эпителий — группы клеток, плотно прилежащих друг к другу боковыми стенками и имеющих апикальную и базальную поверхности. Как мезенхима, так и эпителии могут быть образованы из любого из трех зародышевых листков. Клетки мезенхимного типа наиболее подвижны, так как не образуют между собой стойких контактов. Наиболее яркий пример миграции мезенхимных клеток связан с нервным гребнем. При смыкании нервной трубки клетки нервных валиков выходят из ее состава и располагаются между ее дорсальной частью и эктодермой (см. рис. 7.9).
Рис. 8.1. Пути миграции клеток нервного гребня: а—поперечный срез зародыша, б—производные клеток нервного гребня у взрослого организма; 1—нервный гребень, 2—узел спинного корешка, 3—пигментные клетки, 4—симпатический узел, 5—развивающийся надпочечник, 6—нервное сплетение в стенке кишки, 7— клетка шванновской оболочки, 8—униполярный чувствительный нейрон, 9—клетка-спутник, 10—мультиполярный нейрон симпатического узла, 11—хромаффинная клетка в мозговом веществе надпочечника, 12—превертебральное сплетение, 13— парасимпатическое сплетение в кишке; стрелками показано направление миграции клеток нервного гребня Затем они мигрируют в разных направлениях, проявляя очень широкие формообразовательные потенции. Группа клеток нервного гребня в туловищной части зародыша мигрирует в эктодерму и там превращается в первичные пигментные клетки — меланоциты. Другие, двигаясь в центральном направлении, образуют нейроны спинальных ганглиев, еще дальше — ганглиев симпатической и парасимпатической систем. Третьи — превращаются в клетки шванновских оболочек нервов, четвертые — в хромаффинные клетки мозгового вещества надпочечников. Схематическое изображение этих миграций показано на рис. 8.1. Вообще клетки нервного гребня туловищного отдела дифференцируются в зависимости от того, куда попадут. Клетки нервного гребня в головной части зародыша мигрируют в сторону лица, превращаясь в хрящевые, мышечные, соединительнотканные. Они строят хрящи висцерального скелета, мышцы кожи, соединительную ткань лица, языка и нижней челюсти, входят в состав аденогипофиза, паращитовидных желез и мякоти зуба. Если клетки головного отдела нервного гребня пересадить в туловищную область, то они все равно дифференцируются затем в хрящевые, мышечные и соединительнотканные. Еще одним убедительным примером выраженных миграций является перемещение первичных половых клеток из желточной энтодермы в зачаток половой железы. Нарушение, миграции клеток в ходе эмбриогенеза приводит к недоразвитию органов или к их гетеротопиям, изменениям нормальной локализации. То и другое представляет собой врожденные пороки развития. Примеры пороков развития, связанных с нарушениями миграции клеток, известны, в частности, в отношении конечного мозга. Если нарушается миграция нейробластов, то возникают островки серого вещества в белом веществе, при этом клетки утрачивают способность к дифференцировке. Более выраженные изменения миграции приводят к микрогирии и полигирии (большое число мелких и аномально расположенных извилин больших полушарий), либо, наоборот, к макрогирии (утолщение основных извилин), либо к агирии (гладкий мозг, отсутствие извилин и борозд больших полушарий). Все эти изменения сопровождаются нарушением цитоархитектоники и послойного строения коры, гетеротопиями нервных клеток в белом веществе. Подобные пороки развития отмечены и в мозжечке. Существуют гипотезы о дистантных воздействиях на клетки на основе хемотаксиса и о контактных воздействиях. Мезенхимные клетки способны к амебоидным движениям. Движение их по типу хемотаксиса показано для некоторых видов специализированных клеток (гоноциты, сперматозоиды, некоторые клетки крови). Для эмбриональных клеток многоклеточных животных достоверных случаев хемотаксиса не обнаружено. Контактные взаимодействия более распространены. Они представляют собой взаимодействие клеток со структурированным субстратом. Клетки ощущают микроструктуру субстрата и движутся вдоль волокон, как это происходит при помещении фибробластов на каплю кровяной плазмы, натянутой в виде пленки между сторонами стеклянного треугольника. На неструктурированной подложке, например на ненатянутой пленке, фибробласты перемещаются беспорядочно. Хорошо известно также, что нервные волокна проходят часто длинный путь по различным тканям от нервного центра к рецептору или эффектору и «узнают» их. Скорее всего нервные окончания растут по микроструктурам субстрата, как они делают это и в культуре тканей. Субстратом для движения может быть и соседняя клетка, если оболочка ее натянута. А так как поверхность направленно движущейся клетки сама вытягивается, то и она может служить субстратом для движения и поляризации следующей за ней клетки.
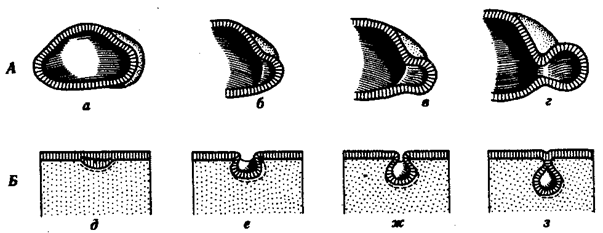
Рис. 8.2. Перемещение клеточных пластов. А —путем выпячивания на примере образования глазного пузырька; Б—путем впячивания на примере образования слухового пузырька: а — стенка переднего мозга, б —местное ускорение роста, в — выпячивание, г — глазной пузырек, д—ж —углубление ямки, з —отшнуровывание пузырька
Согласованные перемещения пластов эпителиальных клеток также изучаются. К ним относятся изгибы клеточных пластов путем выпячивания или впячивания, отшнуровка, образование утолщений — плакод (рис. 8.2). Таким, образом, несомненно, что для миграции клеток очень важны их способность к амебоидному движению и свойства клеточных мембран. И то, и другое генетически детерминировано, так что и сама миграция клеток находится под генетическим контролем, с одной стороны, и влияниями окружающих клеток и тканей — с другой.
Сортировка клеток
В процессе эмбриогенеза клетки не только активно перемешаются, но и «узнают» друг друга, т.е. образуют скопления и пласты только с определенными клетками. Значительные координированные перемещения клеток характерны для периода гаструляции. Смысл этих перемещений заключается в образовании обособленных друг от друга зародышевых листков с совершенно определенным взаимным расположением. Клетки как бы сортируются в зависимости от свойств, т.е. избирательно. Замечено, что необходимым условием сортировки являются степень подвижности клеток иособенности их мембран. В поздней бластуле амфибий, например, клетки будущей эктодермы обладают тенденцией слипаться друг с другом и распространяться в виде сплошного слоя над мезодермой и энтодермой. Эта тенденция проявляется и в культуре тканей. Клетки мезодермы имеют тенденцию впячиваться в любой находящийся поблизости комок клеток, а клетки энтодермы относительно неподвижны. Эти особенности трудно объяснить, если наблюдать только за нормальным течением развития. Впервые на различия в подвижности и способности к слипанию (адгезии) между клетками, относящимися к разным зародышевым листкам, обратил внимание Г. Гольтфретер в 30-х гг. XX столетия. Затем производили модификации его опытов с самыми разными клетками. Смысл этих опытов состоит в том, что зародыши тритонов или других животных на стадии гаструлы диссо-циируют с помощью фермента трипсина, который разрушает материал, соединяющий клетки друг с другом. Диссоциированные (дезагрегированные) клетки тщательно перемешивают и затем создают такие условия, чтобы клетки могли свободно перемещаться и воссоединяться друг с другом. Сначала клетки представляют собой беспорядочную смесь, затем клетки эктодермы, мезодермы и энтодермы разделяются (сегрегируют) и собираются в отдельные группы, каждая из которых представляет собой клеточный агрегат из однородных клеток. Образуются снова зародышевые листки, располагающиеся иногда даже в обычном для них порядке (рис. 8.3).
Рис. 8.3. Адгезия клеток зародышевых листков:
а—смесь диссоциированных клеток гаструлы амфибий, б—клетки эктодермы, мезодермы и энтодермы, группирующиеся послойно путем адгезии
Подобную агрегацию клеток зародышевого листка с себе подобными можно объяснить способностью к избирательному слипанию клеток одного типа между собой. Одновременно это является проявлением ранней дифференцировки клеток на стадии гаструлы. Существует ряд гипотез, объясняющих избирательную сортировку клеток. Возможно, что контакты между подобными клетками сильнее, чем между чужеродными клетками, из-за различий в поверхностном заряде их мембран. Обнаружено, что поверхностный заряд клеток мезодермы ниже, чем клеток экю- или энтодермы, благодаря чему клетки мезодермы легче деформируются и втягиваются в бластопор в начале гаструляции. По другой гипотезе, контактные взаимодействия между одинаковыми клетками основываются на антигенных свойствах их мембран. Избирательная адгезия клеток определенного зародышевого листка друг с другом является необходимым условием нормального развития. Это показано путем сопоставления двух процессов: течения гаструляции в естественных условиях и способности клеток определенного зародышевого листка агрегировать в комплекс с себе подобными. Показано, что если у межвидовых гибридов зародышей амфибий нарушена гаструляция, то диссоциированные в опыте клетки зародышевых листков таких гибридов не способны к агрегации с себе подобными, если же в некоторых случаях гибридам удавалось образовать гаструлу, то и в эксперименте диссоциированные клетки зародышевых листков были способны к реагрегации. По-видимому, в обеспечении сортировки клеток важное место принадлежит генетическим механизмам. Одним из примером потери клетками способности к избирательной сортировке и слипанию является их беспорядочное поведение в злокачественной опухоли. Таким образом, сортировка клеток и их избирательная адгезия наряду с другими клеточными процессами играет важную роль в морфогенезе развивающегося зародыша и одновременно подвержена многоуровневым регуляционным воздействиям (генетическим, межклеточным, онтогенетическим), отражая целостность организма как системы. Гибель клеток
В развитии зародышей наряду с размножением клеток важную роль играют процессы гибели клеток. В настоящее время различают два принципиально различных типа клеточной гибели: апоптоз (в переводе с греческого «отпадающий») и некроз. Апоптоз широко распространен и типичен для физиологических условий. Наряду с описанными выше делением, сортировкой и миграцией клеток, он способствует достижению характерных для определенного биологического вида черт его морфофункциональной организации. Следовательно, апоптоз является естественным, эволюционно обусловленным и генетически контролируемым механизмом морфогенеза. Некроз клеток возникает в нефизиологических условиях, например, в связи с действием неблагоприятных факторов, таких, как стойкое кислородное голодание, разного рода токсины и другие. Некроз обычно сопровождается воспалением и является патологическим процессом. Указанные воззрения в отношении апоптоза и некроза сформировались за последние три десятилетия и поэтому еще нередко некоторые физиологические процессы клеточной гибели описываются термином «некроз» вместо термина «апоптоз». Наиболее яркие примеры разрушения клеток и органов относятся к постэмбриональным стадиям метаморфоза земноводных и насекомых. У головастиков резорбируются (рассасываются) хвост, кишечник и жаберные крышки, у личинок насекомых разрушается большинство внутренних органов. В ходе эмбрионального развития высших позвоночных и человека также имеют место процессы дегенерации органов, которые вначале закладываются, а затем исчезают. У особей женского пола дегенерируют вольфовы протоки, у особей мужского пола—мюллеровы протоки, что является, по-видимому, результатом влияния половых гормонов. У эмбриона человека вначале закладываются ребра у 7-го шейного позвонка и 9—10 хвостовых позвонков, затем они обычно исчезают, так что шейные позвонки, как правило, ребер не несут и в копчике остается 4—5 позвонков (рис. 8.4).
Рис. 8.4. Область клоаки и хвоста у зародышей человека. А —в конце 4-й недели; Б—на 8-й неделе: 1—ректальная область клоаки, 2—уроректальная складка, 3—область мочеполового синуса в клоаке, 4—хорда, 5—нервная трубка, 6—половой бугорок, 7—аллантоис, 8—желточный стебелек, 9—прямая кишка, 10—уроректальная перегородка, 11—мочеполовой синус, 12—мочевой пузырь Немалое значение принадлежит процессам гибели клеток при образовании полостей тела или сосудов (так называемая кавитация), имеющих вначале вид тяжей без просвета. В центральной нервной системе сначала образуется больше нервных клеток, чем затем сохраняется, так как часть нейронов, не установивших связи со своими мишенями, погибает. Наиболее изучены процессы гибели клеток при образовании дефинитивной формы конечностей птиц и млекопитающих. У цыпленка почки конечностей закладываются в виде боковых утолщений соматоплевры примерно на 55-м часу развития. Они вырастают из тела зародыша в виде выступов, покрытых эктодермой и заполненных мезодермальной тканью. По мере их роста начинают проступать контуры конечностей. Процесс формирования контуров сопровождается отмиранием клеток в ряде мезодермальных участков почки конечности. Участки будущей гибели клеток выявлены с помощью прижизненных красителей. Так называемая задняя некротическая зона (ЗНЗ) обеспечивает формирование контуров проксимальных областей конечностей. Максимальной протяженности ЗНЗ достигает к 96-му часу развития. К этому времени гибнет 1500—2000 клеток, поглощаемых примерно 150 макрофагами. Другие клетки мезодермы, примыкающие к зоне некроза, не погибают. Группа клеток будущей ЗНЗ, пересаженная со своего местонахождения на боковую поверхность тела зародыша за 40 ч до начала ожидаемого некроза, все равно погибает в те же сроки, что и на своем обычном месте. В данном примере проявляется генетическая запрограммированность гибели определенных клеток, которую можно образно назвать «внутренними часами смерти». Если клетки ЗНЗ пересадить на дорсальную поверхность почки крыла, то ее клетки не погибнут. Следовательно, некоторые другие клетки способны предотвратить гибель клеток ЗНЗ. Такой способностью обладают клетки мезодермы из дорсальной ненекротической области крыла. Показано, что контакт клеток ЗНЗ с «защищающими» их от гибели клетками дорсальной мезодермы крыла должен продолжаться не менее 6 сут., после чего они могут уже продолжать существование и без контакта с этими клетками. Таким образом, гибель клеток контролируется не только генетически, но и на уровне клеточных взаимодействий. Очевидно, что в описанных событиях мы наблюдаем апоптоз, несмотря на сохранение в названиях гибнущих зонах — «некротические». Интересно сравнение влияния условий на процессы гибели клеток в дистальной части лапок у цыпленка и утенка. Как известно, у утят гибель клеток в дистальной части лапки невелика, вследствие чего у них между вторым, третьим и четвертым пальцами образуются перепонки. Когда создавали химеры, соединяя мезодерму из почки ноги утки с эктодермой цыпленка и наоборот, подсаживая затем химерные почки конечностей на боковую поверхность куриного зародыша, в обоих случаях получали развитие по типу утиной конечности, т.е. с перепонкой между пальцами (рис. 8.5).
Рис. 8.5. Развитие задней конечности цыпленка (А) и утенка (Б); мутация talpid (В) и опыты по пересадке мезодермы и эктодермы в почках конечностей {Г): 1—мезодерма почки конечности, 2—апикальный эктодермальный гребень, 3—участки некроза между пальцами у зародыша цыпленка, 4—химерные почки конечностей, 5—мезодерма утки, 6—эктодерма цыпленка, 7—мезодерма цыпленка, 8—эктодерма утки Таким образом, мезодерма цыпленка, обладая способностью к гибели при нормальном развитии, в присутствии эктодермы утки не погибает, т.е. изменяет свои свойства. Приведенный опыт подтверждает значение межклеточных регуляторных механизмов в развитии. Генетический контроль клеточной гибели прослеживается также на примере мутаций. Анализ мутации под названием ta (talpid) у цыпленка показывает, что в конечности отсутствуют передняя и задняя некротические зоны, в результате чего не происходит формирования контуров, сужающих конечность, и лапка получается очень широкой. Кроме того, у таких мутантов отсутствует область апоптоза под названием темного пятна, в результате чего не происходит разделения большой и малой берцовых костей. Неразделение костей голени и отсутствие их сужения сочетаются с костной полидактилией (многопалостью) и синдактилией (сращением) мягких тканей в ноге. Конечность имеет широкий и веслообразный вид с 6—7 пальцами. Это аутосомно-рецессивная мутация. Аналогичные пороки описаны у человека. Существуют также мутации, расширяющие зоны апоптоза. У дрозофилы мутация Bar сильно уменьшает глаза, а мутация vestigial уменьшает крылья за счет увеличения участков гибели клеток. Описаны подобные мутации и у кур. Новейшим примером генетического контроля апоптоза является открытие гена р53. Белок, контролируемый этим геном, обладает способностью при определенных условиях блокировать клеточное деление и запускать механизм апоптоза. Мутационные изменения и дефектность функции этого гена или регулирующих его активность нуклеотидных последовательностей ДИК встречаются в опухолевых клетках и обнаружены, по данным разных исследователей, у 55—70% раковых больных. Таким образом очевидно, что избирательная гибель клеток, не менее важна для морфогенеза, чем другие клеточные процессы. 'Различные тонкие по замыслу опыты позволяют установить, что гибель клеток имеет три уровня регуляции: генетический контроль, межклеточные взаимодействия и организменный уровень.
Дифференцировка клеток
Дифференцировка — это процесс, в результате которого клетка становится специализированной, т.е. приобретает химические, морфологические и функциональные особенности. В самом узком смысле это изменения, происходящие в клетке на протяжении одного, нередко терминального, клеточного цикла, когда начинается синтез главных, специфических для данного клеточного типа, функциональных белков. Примером может служить Дифференцировка клеток эпидермиса кожи человека, при которой в клетках, перемещающихся из базального в шиповатый и затем последовательно в другие, более поверхностные слои, происходит накопление кератогиалина, превращающегося в клетках блестящего слоя в элеидин, а затем в роговом слое — в кератин. При этом изменяются форма клеток, строение клеточных мембран и набор органоидов. На самом деле дифференцируется не одна клетка, а группа сходных клеток. Примеров можно привести множество, так как в организме человека насчитывают порядка 220 различных типов клеток. Фибробласты синтезируют коллаген, миобласты — миозин, клетки эпителия пищеварительного тракта — пепсин и трипсин. 338 В более широком смысле под дифференцировкой понимают постепенное (на протяжении нескольких клеточных циклов) возникновение все больших различий и направлений специализации между клетками, происшедшими из более или менее однородных клеток одного исходного зачатка. Этот процесс непременно сопровождают морфогенетические преобразования, т.е. возникновение и дальнейшее развитие зачатков определенных органов в дефинитивные органы. Первые химические и морфогенетические различия между клетками, обусловливаемые самим ходом эмбриогенеза, обнаруживаются в период гаструляции. Зародышевые листки и их производные являются примером ранней дифференцировки, приводящей к ограничению потенций клеток зародыша. На схеме 8.1 представлен пример дифференцировки мезодермы (по В. В. Яглову, в упрощенном виде). Популярное:
|
Последнее изменение этой страницы: 2016-04-11; Просмотров: 969; Нарушение авторского права страницы