|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ФИЗИКО-ХИМИЧЕСКИЕ ИЗМЕНЕНИЯ МЫШЕЧНОЙ ТКАНИ ПРИ АВТОЛИЗЕ
В результате интенсивного накопления кислых продуктов в начальных стадиях автолиза происходит насыщение буферных систем мышечной ткани. Значительное накопление фосфорной, молочной, пировиноградной кислот является причиной почти полного разрушения бикарбонат-ного буфера и выделения углекислоты уже в первые часы после убоя животного. Наиболее важным является буферное действие белков. Однако интенсивное накопление кислот приводит к сдвигу рН в кислую зону сначала до 6, 2—6, 0, а затем до 5, 8 и 5, 6. Характерное изменение свободной кислотности мышечной ткани в значительной мере обусловлено особенностями изменений белков. Так, в период мышечного окоченения уменьшение буферной способности белков является следствием уплотнения их структур и результатом интенсивных агрегационных взаимодействий. При разрешении окоченения и протеолитической деструкции прояв-ляется более высокое связывание белками водородных ионов и убыль их в среде (повышение рН). Этому же спо-собствует (повышает буферную способность) накопление веществ небелковой природы (продуктов автолиза). В автолизирующих мышцах накапливающиеся активные кислоты вытесняют катионы из различных комплексов, в которых они были связаны при жизни животного. Воздействие электролитов и автолитический распад липо-протеидов приводят к нарушению целостности внутриклеточных и клеточных мембран, что обусловливает перераспределение ионов и жидкости. Некоторое количество ионов калия диффундирует из клеток во внеклеточную жидкость, а ионы натрия, наоборот, поступают из внеклеточной жидкости в клетки. Другие катионы, как и анионы, свободно диффундируют через клеточную мембрану, вследствие чего рН клеток и внеклеточной жидкости выравнивается. В результате перераспределения катионов и анионов изменяются физико-химические свойства белков, что является одной из причин изменения их растворимости и гидратации, изменяется активность ферментов. ИЗМЕНЕНИЕ РАСТВОРИМОСТИ БЕЛКОВ В начальных стадиях автолиза мышечной ткани происходит уменьшение растворимости мышечных белков, а затем после достижения определенного минимума повышение их экстрагируемости. В этот период автолиза мышц млекопитающих и птиц для всех белков характерны связанные с изменением зарядов конформационные сдвиги, стимулирующие агрегационные взаимодействия белков. Характер этих изменений для однотипных белков разных мышц неодинаков и соответствует интенсивности накопления кислот (продуктов автолиза небелковой природы). Поэтому и извлекаемость однотипных белков разных мышц при одинаковых условиях автолиза различна. Белки саркоплазмы. В начальный период автолиза наибольшее снижение экстрагируемости белков саркоплазмы выявляется у мышц, в которых интенсивно происходит накопление продуктов автолиза небелковой при- роды (фосфорной, молочной, пировиноградной, кетокис-дот и др.). Действие этих продуктов обусловливает изменения, способствующие большому межмолекулярному взаимодействию белков (рис. 16). Взаимодействие белковых частиц в этот период, по-видимому, осуществляется в основном за счет электростатических сил, так как извлечение белков саркоплазмы буферными растворами физиологической концентрации с высокой диэлектрической постоянной повышает их экстрагируемость.
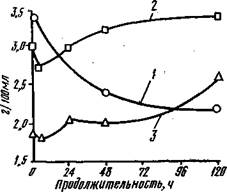
Рис. 16. Суммарная экстрагируемость белков саркоплазмы при автолизе (4° С) мышц: / — крупного рогатого скота; 2, 3 — соответственно белых и красных мышц кур. Извлекаемость разных фракций белков саркоплазмы при автолизе уменьшается неодинаково. Наиболее заметно уменьшается экстрагируемость белков фракций глобулина X (рис. 17, фракция //) и миоальбуминов (рис. 17, фракция /). Свойства белков миогеновой группы (рис. 17, фракции ///, IV и IV д) изменяются меньше. У этих белков хорошо сохраняется растворимость и ферментативная активность. У высокомолекулярных белков (фракция V) изменения незначительны. После достижения минимума (по времени для различ
Рис. 17. Изменения экстрагируемое™ фракций (I—V) белков саркоплазмы: J — дорзальных мышц крупного рогатого скота; 2, 3 — соответственно белых и красных мышц кур (IV д — дополнительная фракция миогеновой группы белков). Белки миофибрилл. В начальных стадиях автолиза мышц уменьшается растворимость миозина в результате образования менее растворимого комплекса актомиозина. Актин довольно прочно удерживается в структуре миофибрилл, поэтому и связанный с ним миозин не извлекается без воздействия веществ, деполймери-зующих актомиозиновый комплекс. Уменьшение экстрагируемости миозина в автолизирующих мышцах в этот период происходит за счет снижения концентрации АТФ (добавление к измельченным мышцам АТФ или пирофосфата восстанавливает растворимость миозина). В процессе окоченения белки миофибрилл претерпевают конформацион-ные изменения, способствующие интенсивным аг-регационным взаимодействиям. Понижение экстрагируемости миофибрил-лярных белков продолжается до определенного периода. Например, в мышцах крупного рогатого скота до 24—48 ч автолиза, а для мышц птиц этот срок значительно меньше. Затем происходит повышение их растворимости за счет как диссоциации актомиозинового комплекса, так и ослабления агрегаци-онных взаимодействий вследствие перераспределения зарядов. Одной из причин повышения экстрагируемости является также ограниченная протеолитическая деструкция миофибриллярных белков. ИЗМЕНЕНИЕ ВОДОСВЯЗЫВАЮЩЕЙ СПОСОБНОСТИ МЫШЕЧНОЙ ТКАНИ
уменьшения водосвязыва- 40 ющей способности согласуется с особенностями изменений миофибрилляр-ных белков, а также особенностями их агрегаци-онных взаимодействий. Выявляется определен ткани оказывает характер изменений и остальных белков мышечной ткани. В начальных стадиях автолиза в ре. зультате подкисления большая часть белков мышечной ткани переходит в изоэлектрическое состояние, что способствует лучшей агрегации белков и уменьшению гид. ратационной способности. Поэтому как повышение, так и понижение рН среды от изоэлектрического состояния белков (рН 5, 5) приводит к повышению гидратации мышц. С началом разрешения окоченения происходит постепенное повышение водосвязывающей способности мышц. При повышении водосвязывающей способности четко выявляется повышение активности гидрофильных групп в белках мышц, «разрыхление» белковых структур. При этом активность свободных кислотных групп в белках мышц выявляется в большей степени, чем основных. Это объясняется тем, что белки мышц содержат больше ди-карбоновых аминокислот (глютаминовой и аспарагино-вой), чем диаминокислот (лизин и аргинин). Таким образом, белки мышц являются главными связывающими воду соединениями и в первую очередь от их состояния, изменения их структуры и состояния гидрофильных групп зависит водосвязывающая способность мышечной ткани. ИЗМЕНЕНИЕ АКТИВНОСТИ ФЕРМЕНТОВ В процессе автолиза мышечной ткани характер автолити-ческих превращений меняется во времени, изменяется соотношение скоростей биохимических реакций. Выход ферментов из структур при автолизе мышечной ткани повышает объем ферментативных превращений. В полиферментных системах реакции легко протекают и тогда, когда ферменты отделены друг от друга, а доступ к ним осуществляется путем свободной диффузии. В начальных стадиях автолиза наиболее активными оказались ферменты гликолитической системы, а-амила-за, а-олигоглюкозидазы, миозиновая АТФ-аза; активны ряд оксидоредуктаз — сукцинатдегидрогеназа, цитохро-моксидаза и др. По мере накопления продуктов автолиза небелковой природы и подкисления в клетках мышечной ткани происходят распад липопротеидных оболочек лизосом, вьь сВобождение катепсинов, гликозидаз, кислой фосфатазы, рйбонуклеазы и дезоксирибонуклеазы и других гидроли-^лческих ферментов, а также их активация. Почти все эТи ферменты наиболее эффективны в слабокислой среде 0 поэтому известны под общим названием кислых гидро- лаз. Активными оказались и ферменты, связанные с митохондриями. По мере разрушения в процессе автолиза этих органелл и выхода ферментов активность их проявлялась в достаточной степени. Так, значительную активность проявили аминоферазы и декарбоксилазы целого-ряда аминокислот. Однако в дальнейшем по мере развития автолитиче-ских превращений в результате накопления продуктов автолиза, высвобождения и перераспределения анионов и; катионов и взаимодействия их с ферментами происходит снижение активности многих энзимов. Это является результатом химических взаимодействий с отдельными функциональными группами активного центра фермента,, специфического взаимодействия с отдельными участками апофермента, неспецифической адсорбции на поверхности белка, а также взаимодействия стеми группами, которые поддерживают специфическую конформацию молекулы белка — фермента. Кроме того, для многих ферментов, в том числе и для миозиновой АТФ-азы, потеря активности может быть и следствием протеолитической деструкции самого фермента. ПРОТЕОЛИТИЧЕСКИЕ ПРЕВРАЩЕНИЯ В соответствии с особенностями метаболизма разные виды мышечной ткани отличаются различной концентрацией катепсинов. Наряду с этим в соответствии с функциональными особенностями разные мышцы одного и того же животного также отличаются различной концентрацией этих ферментов. Как указывалось ранее, катепсины заключены в лизосомы. В начальных стадиях автолиза по-мере распада одинарной липопротеидной оболочки лизосом катепсины выходят из ограничивающих их структур в Цитоплазму, в результате чего и происходит нарастание свободной протеолитической активности. Возможно, в силу различных способов ограничения начальная свободная активность, степень нарастания протеолитической актив- ности у разных видов автолизирующих мышц неодинако. вы. У мышц с высоким прижизненным метаболизмом в начальных стадиях автолиза протеолитическая активность нарастает быстрее, что и обусловливает соответствующие интенсивные протеолитические превращения. Высвобождение катепсинов и проявление их активности находятся в прямой связи с течением гликолитических и других автолитических превращений компонентов небелковой природы, связанных с накоплением электролитов. Развитие этих процессов способствует разрушению липопротеидной оболочки лизосом. Кроме того, подкисле-ние ткани является также важнейшим фактором для стимуляции действия катепсинов, нарушающих целостность оболочки лизосом изнутри. Однако длительное воздействие электролитов приводит к снижению протеолитическои активности. Опыты по введению в измельченную мышечную ткань хлористого натрия показали инактивацию катепсинов в этих условиях. Вместе с тем воздействие указанных факторов является неравномерным, поэтому автолизирующая мышечная ткань на различных сроках автолиза обладает определенной остаточной (и для многих видов мышц — значительной) протеолитическои активностью. Величина остаточной протеолитическои активности находится в прямой зависимости от концентрации катепсинов и в значительной мере в обратной зависимости от количества накопившихся продуктов автолиза небелковой природы. Ограниченному протеолизу в соответствии с локализацией в первую очередь подвергаются белки саркоплазмы под воздействием высвобождающихся тканевых эндопеп-тидаз. Это приводит к возникновению в белках N-концевых и С-концевых остатков аминокислот, некоторой деструкции белковых макромолекул. Такие превращения обусловливают повышение общей растворимости и элект-рофоретической подвижности белков саркоплазмы. С увеличением продолжительности автолиза электрофоретиче-ская картина в целом изменяется, появляются группы модифицированных белковых фракций, подвижности которых варьируют с небольшой дисперсией. Миофибриллярные белки также подвергаются ограниченной протеолитическои деструкции. В белках фракции миозина накапливаются N-концевые группы аминокислот. Среди N-концевых остатков большинство составляют ^{отаминовая и аспарагиновая кислоты, лейцин, в меньшем количестве выявляются серии, треонин, тирозин и ва-лйН- В результате такого ограниченного протеолиза миофибриллярные белки становятся более растворимыми, п0вышается доступность их сульфгидрильных, дисуль-флдных и других химических групп. В результате ограниченного протеолиза всех белков повышается реактивность кислых и основных групп в белках мышц, водосвязывающая способность, пластичность и нежность мышц. Накопление в процессе автолиза относительно значительных количеств свободных аминокислот по сравнению с исходным их содержанием свидетельствует о проявлении наряду с эндопептидазами также достаточной активности различных тканевых экзопептидаз, атакующих концевые пептидные связи, а также трипептидаз и дипепти-даз. Преимущественное накопление в процессе автолиза мышц а-аланина, глицина, глютаминовой кислоты, вали-на, фенилаланина, тирозина, треонина, лейцина и изолей-цина отражает специфическое совместное воздействие на белки и пептиды тканевых эндопептидаз и экзопептидаз. Накопление р-аланина является также следствием ферментативного гидролиза специфических дипептидов. Наряду с этим в автолизирующей мышечной ткани накапливается орнитин. Начальное содержание этой аминокислоты невелико. Однако в процессе автолиза мышц происходит рост ее содержания, который, очевидно, связан с распадом аргинина, поскольку одновременно происходит и накопление мочевины. В процессе автолиза мышц наряду с накоплением многие аминокислоты подвергаются различным превращениям. Такие аминокислоты, как гистидин, тирозин, глюта-миновая кислота, триптофан, заметно декарбоксилируют-ся. То же наблюдается для 5-окситриптофана и цистеино-вой кислоты — продуктов обмена триптофана и цистеина. В результате этого автолизированные мышцы содержат заметное количество циклических аминов (в порядке возрастания): гистамина, тирамина, триптамина. Накапливается в больших количествах таурин. Накопление аминов при стерильном автолизе мышц является не только следствием декарбоксилирования аминокислот, но и результатом торможения окислительного дезаминирования
4—454 аминов, поскольку отсутствует поступление кислорода в клетки мышц. В процессе автолиза мышц происходит сравнительно небольшой объем переаминирования аминокислот и накопление их кетоаналогов. К числу аминокислот, заметно вступающих в реакции переаминирования, относятся: а-аланин, глютаминовая и аспарагиновая кислоты. Наибольшим количественным превращениям в процессе автолиза мышц подвергается глютаминовая кислота. Помимо декарбоксилирования и участия в процессах переаминирования, значительная часть глютаминовой кислоты, по-видимому, расходуется в реакциях биохимического связывания аммиака и образования ее амида — глютамина. ИЗМЕНЕНИЯ УЛЬТРАСТРУКТУР В ПРОЦЕССЕ АВТОЛИЗА Автолиз мышц сопровождается изменениями структур ор-ганелл, миофибрилл, саркоплазматического ретикулума, сарколеммы и других образований мышечного волокна. На начальных стадиях автолиза начинает изменяться форма частиц гликогена в мышечном волокне. К 12 ч при низких положительных температурах резко уменьшается содержание гликогена, а к исходу 24 ч гистохимически гликоген уже не обнаруживается. К концу первых суток автолиза при низких положи При длительном автолизе в зависимости от степени деструктивных изменений мышечные волокна теряют способность сокращаться при добавлении АТФ. По уменьше- вию контр акционной способности можно судить о степени деструктивных изменений миофибриллярной структуры. Добавление АТФ к автолизированным мышечным волок-лам вызывает растворение разрушенных микроструктур, после чего остается только «остов» миофибриллярных структур. Изменение ультраструктур в автолизирующих мышцах следует рассматривать не только как следствие ферментативных гидролитических превращений, но и как результат физико-химических и коллоидно-химических изменений белков мышечного волокна. Такие изменения интенсивно проявляются в процессах окоченения и расслабления мышечной ткани. Изучение сложных и многообразных превращений при окоченении и расслаблении мышц показало, что уже в начальных стадиях этих процессов происходит молекулярная перестройка белков ак-томиозинового комплекса, а это ведет к перестройке субмолекулярных структур. Значительные изменения ультраструктур в мышцах выявляются в процессах холодильной обработки и посола. Высокие агрегационные взаимодействия белков при замораживании и хранении в мороженом виде мышц обусловливают и изменения субмолекулярных структур. Изменение ультраструктур вызывает и растворяющее действие гипертонических растворов при размораживании и посоле.
4* Глава II. БИОХИМИЯ КРОВИ БИОХИМИЧЕСКИЕ ФУНКЦИИ, СВОЙСТВА И СОСТАВ КРОВИ БИОХИМИЧЕСКИЕ ФУНКЦИИ КРОВИ Кровь (жидкая ткань животного организма) является внутренней средой организма, которая объединяет между собой органы и ткани и выполняет дыхательные, питательные, выделительные, регуляторные и защитные функции. Она доставляет к органам и тканям питательные вещества, поступающие из внешней среды через пищеварительный тракт, и в нее выделяются из тканей различные продукты обмена. Артериальная кровь переносит кислород, необходимый для тканевого дыхания. В кровь выделяются гормоны, т. е. она способствует гормональной регуляции. Кроме того, регуляторные функции крови обусловливаются постоянным обогащением ее ферментами и другими биологически активными веществами. Регуляторные функции заключаются также в поддержании постоянства осмотического давления, активной реакции среды и температуры тела. Важнейшей функцией крови является транспортировка конечных продуктов тканевого обмена (например С02, мочевины, мочевой кислоты, аммонийных солей и других азотистых веществ, а также избытка минеральных солей) к органам выделения. Кровь выполняет защитные функции организма, участвуя в борьбе организма с многими видами заболеваний. При попадании в кровь или ткани животного организма инородных высокомолекулярных веществ (белки или полисахариды), называемых антигенами, образуются антитела — белки, специфически реагирующие с исходными антигенами. Антитела вырабатываются плазматически- яи клетками, которые сходны с лимфоцитами и находятся в селезенке, лимфатических узлах, стенке пищеварительного тракта, печени и в других органах. Антитела способствуют агглютинации (лат. agglutinare — приклеивать) бактерий и связыванию антигенов. Обнаруживают антитела по способности соединяться с исходным антигеном. Они исключительно специфичны и не реагируют даже с молекулами, сходными по строению с антигеном. Защитные функции крови определяются также лейкоцитами, которые богаты протеолитическими и липолити-ческими ферментами, способствующими быстрому распаду и перевариванию различных микробных тел при фагоцитозе. МОРФОЛОГИЧЕСКАЯ ХАРАКТЕРИСТИКА КРОВИ Кровь состоит из жидкой части — плазмы — и взвешенных в ней форменных элементов. К форменным элементам относятся: эритроциты (красные кровяные тельца) — специфические клетки крови, безъядерные у большинства животных или с ядрами, например, у птиц, амфибий, рептилий; лейкоциты (белые кровяные тельца) — лимфоциты, моноциты, нейтрофилы, эозинофилы и базофилы; в этих клетках имеются ядра, однако циркулирующие лейкоциты не делятся; тромбоциты (кровяные пластинки, бляшки) — продукты фрагментации (распада) особых гигантских клеток костного мозга — мегакариоцитов. В крови разных видов животных содержание форменных элементов неодинаково. Количество их в среднем составляет у крупного рогатого скота 33%, у мелкого 28%, свиней 43, 6%, лошадей около 40% от массы крови. Общее количество крови у различных видов животных также неодинаково. Так, у крупного и мелкого рогатого скота оно составляет 7, 6—8, 3%, свиней 4, 5—6, 0%, домашней птицы 8, 1%, лошадей 9, 8%, кроликов 5, 5—6, 2% к массе животных. ХИМИЧЕСКИЙ СОСТАВ И ФИЗИКО-ХИМИЧЕСКИЕ СВОЙСТВА КРОВИ Химический состав крови у животных одного вида в норме постоянен, а у животных разных видов имеются некоторые колебания в содержании компонентов. Химический состав цельной крови убойных животных (средние данные) приведен в табл. 4. ТАБЛИЦА 4 Содержание составных частей (в г) в 1000 г цельной крови Состав крови быка
Вода.................. Сухой остаток... В том числе:
гемоглобин.. другие белки
сахар............ холестерин..
лецитин....
жир........... жирные кислоты
натрий.............. калий............. окись железа кальций....
I Кровь является средой, в которую поступают различные продукты тканевого обмена, кислород, а также питательные вещества. Несмотря на это, кровь большого круга кровообращения характеризуется довольно постоянным составом. Такое динамическое постоянство поддерживается различными механизмами, регулирующими поступление в кровь продуктов питания и выделение из нее продуктов обмена. Благодаря постоянному составу плазмы крови создаются неизменные условия среды, необходимые для существования клеток. Осмотическое давление крови животных обусловливается в основном содержанием в ней неорганических ве- ществ (их молекул и ионов), а также частично белками рлазмы и другими органическими соединениями. Осмотически наиболее активными веществами являются соли NaCl, NaHC03, Na2HP04, СаСЬ, KC1, при диссоциации которых образуются ионы, ведущие себя как осмотически активные частицы. Постоянство осмотического давления крови обеспечивается деятельностью регуляторных механизмов, способствующих выделению из организма избытка как воды, так и осмотически активных веществ. Осмотическое давление выражают величиной депрессии (А), характеризующей температуру понижения замерзания крови по сравнению с температурой замерзания чистой воды. Депрессия крови убойных животных составляет: коровы 0, 611, овцы и свиньи 0, 618, лошади 0, 558, кролика 0, 595. Ряд жизненных функций клеток, в частности сохранение ими определенной формы, способность воспринимать извне и отдавать во внешнюю среду вещества, зависит от осмотического давления клеточного сока и омывающей клетки жидкости, т. е. лимфы и связанной с ней плазмы крови. Осмотическое давление белков плазмы и других коллоидов называется коллоидно-осмотическим, или онкоти-ческим, давлением. Оно примерно составляет около 0, 004—0, 006 общей величины осмотического давления крови. Осмотическая (онкотическая) активность белков плазмы неодинакова. Например, сывороточные альбумины обусловливают 3/4 осмотического давления, создаваемого белками, хотя по количеству составляют приблизительно 7г всех белков плазмы/Сравнительно высокая осмотическая активность сывороточных альбуминов является следствием большей их концентрации (в результате меньшей величины молекул) по сравнению с другими белками плазмы. Распределение воды между тканями и кровью при одинаковой концентрации в них солей определяется концентрацией белков в плазме. Если в кровь вводится большое количество физиологического раствора, изотоничного плазме, то концентрация сывороточных белков в крови резко снижается. При этом онкотическое давление в плазме уменьшается, что создает предпосылки для перехода
воды из крови в ткань, поскольку осмотическое давление белков в тканевой жидкости остается прежним. Для крови животных характерно также относительное постоянство концентрации ионов водорода (рН). Реакция крови убойных животных слабощелочная и колеблется обычно в небольших пределах. Кровь рН Коровы......... 7, 36—7, 50 Овцы............. 7, 40—7, 58 Барана........................ 7, 82 Козы........................... 7, 65 Изменение рН крови может быть вызвано рядом причин. В продуктах переваривания, поступающих в кровь, имеются различные кислоты и основания. Кроме того, в продуктах внутриклеточного обмена, поступающих вначале в тканевую жидкость, а затем в кровь, также содержится значительное количество кислот и оснований. Прижизненный сдвиг рН крови в кислую сторону называется ацидозом, в щелочную — алкалозом. Компенсированный сдвиг кислотно-щелочных отношений в сторону алкалоза характерен для травоядных животных, в сторону ацидоза — для плотоядных. ТАБЛИЦА 5
Буферные системы Постоянство активной реакции среды обусловлено наличием в крови буферных систем и непрерывным освобождением ее от конечных продуктов обмена путем экскреции. Главные буферные системы крови приведены в табл. 5. Буферные системы крови отличаются высокой кислотной емкостью. Температура замерзания и электропроводность крови постоянны, что обусловлено в основном постоянным количеством электролитов. Минеральных веществ содержится около 0, 9%. Плотность крови животных разных видов различается незначительно (г/см3).
ПЛАЗМА КРОВИ В плазме крови содержится 90—91% воды и 9—10% плотного остатка. Большую часть плотного остатка составляют белки, а остальную часть — азотистые и без- ТАБЛИЦА 6 Содержание составных частей (в г) в 1000 г плазмы Состав плазмы
Крупного Рогатого Скота
Бикарбонатная.. Фосфатная.... Белковая (плазмы) Гемоглобиновая Оксигемоглобиновая Н2С03 NaHCOg NaH2PQ4 Na2HP04 Н ■ — белок Na — белок н2со3 КНС03 КН2Р04 К2НР04 Н(НЬ) К(НЬ) H(Hb)Q2 К(НЬ)02 Вода.................................... Сухой остаток............... В том числе белки....................... сахар........................... холестерин............... лецитин........................ жир.............................. Жирные кислоты... натрий........................... калий........................... кальций....................... магний............................... хлор............................... Общий фосфор.... в том числе неорганический фосфор... 913, 64 86, 36 72, 50 1, 05 1, 238 1, 675 0, 926 4, 312 0, 255 0, 1194 0, 0446 3, 69 0, 244 0, 085 917, 44 82, 56 67, 50 1, 06 0, 879 1, 709 1, 352 0, 71 4, 303 0, 256 0, 117 0, 041 3, 711 0, 232 0, 073 917, 61 82, 39 67, 741 1, 212 0, 409 1, 426 1, 956 0, 794 4, 251 0, 27 0, 122 0, 0412 3, 627 0, 1972 0, 0524 902, 05 97, 95 84, 24 1, 176 0, 298 1, 720 1, 300 4, 434 0, 263 0, 1113 0, 045 3, 73 0, 240 0, 071
азотистые экстрактивные вещества, липиды, углеводы ^ минеральные вещества. Общий химический состав пладч мы крови убойных животных представлен в табл. 6. БЕЛКИ ПЛАЗМЫ Биохимические и физико-химические свойства белков плазмы Основными фракциями белков плазмы крови являются сывороточные альбумины, сывороточные глобулины д фибриноген. Альбумины и глобулины активно участвуют в обмене веществ, быстро расходуются на нужды организма и так же быстро восстанавливаются. Содержание белковых фракций в плазме разных животных неодинаково (табл. 7). ТАБЛИЦА 7 Содержание (в %) в плазме крови Белковые фракции крупного мелкого
рогатого рогатого
скота скота Фибриноген является главным компонентом системы свертывания крови (см. с. 112). Он нерастворим в воде, но хорошо растворяется в разбавленных растворах нейтральных солей и в щелочах, осаждается сернокислым магнием и хлористым натрием ранее, чем наступает полное насыщение. Фибриноген — белок с молекулярной массой 330 000. Молекулы фибриногена состоят из трех глобулярных структур с диаметром около 6-10~9 м, соединенных между собой более тонкими белковыми тяжами (рис. 19). Молекула фибриногена построена из шести полипептидных цепей, соединенных дисульфидными связями — двух А- и двух В- цепей, несущих у разных животных в качестве N-концевых групп различные кислотные остатки и двух С-цепей — с тирозильными остатками на jsj-конц'е (см. с. 113).
Сывороточные альбумины. Эти белки у различных убойных животных сходны по своим биологическим и физико-химическим свойствам. Сывороточные альбумины, образуемые в печени, поддерживают коллоидно-осмотическое (онкотическое) давление крови. При физиологических значениях рН они заряжены отрицательно, а молекулы их близки по размерам. Внутренняя вязкость сывороточных альбуминов сравнительно невелика, что существенно для кровообращения, так как при этом уменьшается работа, выполняемая сердцем. Сывороточные альбумины участвуют в регуляции кислотно-щелочного равновесия и играют особую роль в транспортировке различных соединений.
Сывороточные альбумины выполняют ответственную роль в переносе метаболитов. Они связываются с углеводами, липидами и другими соединениями. Альбумины — непосредственные акцепторы жирных кислот. Они переносят липиды от жировых депо к местам потребления. Очень велика роль сывороточных альбуминов в переносе углеводов, с которыми они образуют различные глюкопроте-иды. Очень легко возникают комплексы сывороточных альбуминов с различными катионами и анионами, а также с многочисленными продуктами обмена. Наиболее важными являются питательные функции сывороточных альбуминов. В некоторых случаях при очень интенсивном обмене они могут быть единственным источником белкового питания тканей. Альбумины выполняют важную роль в транспортировке и регуляции активности гормонов. По физико-химическим свойствам сывороточные альбумины являются типичными альбуминами: они растворяются в воде и солевых раствор ах среди ей концентрации, причем растворимость их во многих растворителях выще растворимости большинства других компонентов плазмы. В связи с этим методы их выделения основаны на осаждении других компонентов, тогда как альбумины остаются в растворе. Но ввиду того, что сывороточные альбумины легко взаимодействуют с другими белками, а также могут быть связаны с липидами и углеводами, растворимость их в естественных системах может отличаться от растворимости в изолированном состоянии. Очищенный сывороточный альбумин остается гетерогенным, что показано при помощи электрофореза, хроматографии и иммунохимических методов. Способность сывороточного альбумина легко вступать в реакцию с целым рядом веществ обусловлена наличием в его молекуле 55 е-аминогрупп лизина, высокоактивных тиоловых и имидозольных групп, большого количества карбоксильных групп (активность последних при физиологических значениях рН относительно невелика). Популярное:
|
Последнее изменение этой страницы: 2016-06-05; Просмотров: 1355; Нарушение авторского права страницы
 Продолжительность, ч
Продолжительность, ч Непосредственно после убоя животного мышечная ткань
Непосредственно после убоя животного мышечная ткань