
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
В горяче-парном виде из-за высокого значения рН мяса белки находятся в ионизированном состоянии, близ-
НаВухание 180 Щ/40 IOO во 5, 2 W 7, 0 рН Рис. 34. Изменение удержания влаги мясом при набухании и варке (при 70°С): / — мясо парное; 2 — мясо охлажденное. ком к состоянию нативного белка. Благодаря наличию АТФ актин и миозин не связаны между собой. Все это способствует тому, что белки мяса легко гидратируются, набухают, хорошо удерживают влагу и легко переходят в раствор. Поэтому в колбасном производстве парное мясо нет необходимости выдерживать в посоле для улучшения набухания белков. Для набухания и последующего удержания влаги соленым мясом при варке большое значение имеет величина рН. Искусственный сдвиг реакции среды у охлажденного соленого мяса только на 0, 2—0, 3 рН ближе к значению реакции среды парного мяса вызывает резкое увеличение гидратации (рис. 34) и влагоудерживающей
способности мяса. Однако это касается только соленого, а не свежего мяса. На практике для сдвига рН мяса на 0, 2—0, 4 во время измельчения только что посоленного мяса добавляют 0, 2—0, 3% бикарбоната натрия, благодаря чему увеличивается набухание мяса, его адгезия и влагоудержание. На набухание мяса существенно влияет состояние белков в структуре ткани. Так, гидратация мяса увеличивается в результате выделения актомиозина или миозина из структуры ткани. Такой процесс может быть быстро осуществлен путем искусственного введения АТФ или неорганического пирофосфата (пластифицирующий эффект) при измельчении мышечной ткани в присутствии соли и воды и сопровождается резким уменьшением вязкости массы (у охлажденного мяса вязкость с 12, 8—28, 0 снижается до 0, 35—1, 80 Па-с). На практике хорошие результаты получаются при добавлении к мясу препаратов полифосфатов (пирофосфат натрия, гексаметафосфат, триполифосфат и др.). Введение большинства полифосфатов сопровождается изменением состояния белков в результате сдвига рН на 0, 2—0, 3 в сторону нейтральной реакции, а это значительно повышает гидратацию мяса. Кроме того, полифосфаты вызывают диссоциацию актомиозина с освобождением из структуры миозина и увеличением его растворимости. В итоге такое действие фосфатов обусловливает увеличение набухания, адгезии мяса и последующего влаго-удержания при варке мяса. Недавно показано, что при хранении соленого мяса с пирофосфатом при 4° С через 3 суток происходит полное разрушение фосфата: 50% триполифосфата гидролизует-ся сразу после его введения в мясо. Этот распад происходит под действием ферментов — пирофосфатазы (пиро-фосфат-фосфогидролаза) и полифосфатазы. Изменение экстрактивных веществ мяса. Во время посола мяса в рассол диффундируют азотистые и безазотистые экстрактивные вещества мышечной ткани, а также минеральные соединения и витамины. Установлено, что потеря таких азотистых экстрактивных веществ, как креатин, карнозин, карнитин и ряд нуклеотидов, мало зависит от крепости рассола и определяется главным образом соотношением рассола и мяса. При этом возможен переход в рассол до 50% экстрактивных веществ. При мокром посоле из минеральных веществ в рассол диффундируют главным образом фосфаты и калий. В результате этого в мясе уже в первые дни посола быстро уменьшается содержание общего (главным образом неорганического) фосфата (табл. 45). В дальнейшем количество его в мясе существенно не изменяется, но вместе с тем наблюдается значительное уменьшение (почти в 3 раза) количества органических фосфорных соединений, экстрагируемых из мяса солевым раствором (гек-созо- и триозофосфаты, мононуклеотиды). Одновременно с этим наблюдается деструкция органических фосфорных соединений, перешедших в рассол. Возможно, что эти изменения органических фосфорных соединений являются следствием биохимических превращений. ТАБЛИЦА 45
В мясе и в рассоле уменьшается содержание органических фосфорных соединений (фосфолипиды, нуклео-протеиды и др.) и возрастает фракция неорганического фосфора. Изменение процесса автолиза. В результате воздействия компонентов посолочных смесей на белковые вещества и другие соединения мышечной ткани и изменения физико-химических условий среды ферментативные ав-толитические процессы существенно нарушаются, но не прекращаются. Так, при посоле горяче-парного мяса (NaCl и нитрат) тормозятся процессы гликолитического распада. В целом распад гликогена и накопление лакта- та снижается на 20%. Вместе с тем резко усиливается накопление редуцирующих веществ, связанных с интенсивным амилолитическим распадом гликогена в начале посола вследствие активации амилазы хлористым натрием. В результате торможения процессов гликогенолиза реакция среды мышечной ткани сдвигается в кислую сторону менее резко, чем при обычном автолизе. Химические соединения, вводимые в мясо, ингибируют гликолитические и другие ферменты; исключение составляют липолитические. Поваренная соль в концентрации 2—3% оказывает тормозящее действие на ферментную активность миозина. Высокие концентрации нитрата полностью ингибируют фермент. Вследствие ингибирова-ния миозина и торможения распада АТФ уменьшается накопление органического фосфата при автолизе соленого парного мяса. И нитрит, и продукт его превращения ■ — гидроксиамин — ингибируют каталазу. При посоле охлажденного или дефростированного мяса изменения в углеводной системе и в содержании молочной кислоты не существенны, так как главные изменения уже свершились при автолизе до посола. При длительном сухом посоле свиного мяса (окороков) в результате ферментативного протеолиза увеличивается содержание растворимых белков и накапливается остаточный азот. Последовательное изучение изменения содержания свободных аминокислот при продолжительном сухом посоле окороков (24 недели) показало значительное увеличение и суммы (по нингидриновому индексу) и большинства индивидуальных аминокислот. Содержание серина, глютаминовой кислоты, треонина, лейцина, изолейцина, фенилаланина, пролина возросло примерно в 3 раза. Концентрация тирозина, аланина, глицина и гистидина повысилась почти в 2 раза. Уровень метионина, триптофана, таурина, аргинина, карнозина, орнитина, лизина, аспарагиновой кислоты сохранился. Все это подтверждает важную роль катепсинов в процессе созревания ветчины. При мокром посоле мяса процесс протеолиза также происходит. Однако при этом большее значение приобретает развитие специфической микрофлоры, вызывающей своеобразные биохимические превращения его составных частей. Продукты этих превращений влияют на свойства щяса. Отмеченное выше перераспределение ионов и экстрактивных веществ, несомненно, оказывает соответствующее влияние на течение биохимических процессов в системе. Большое влияние на процессы, происходящие при посоле, оказывает рН, а также окислительно-восстановительный потенциал рассола. Величина рН свежего рассола обычно 6, 0—7, 0. Сдвиг реакции среды в более кислую сторону встречается редко, чаще наблюдается нежелательное отклонение рН в щелочную сторону под влиянием микрофлоры, вызывающей распад белков с накоплением оснований, что приводит к порче рассола. В результате развития микрофлоры, окисляющей молочную кислоту, понижается окислительно-восстановительный потенциал и наблюдается нежелательное избыточное разрушение нитратов и нитритов. В случае мокрого посола мяса при низких температурах под действием ферментов микроорганизмов очень незначительная часть белков, перешедших в рассол, распадается до полипептидов, аминокислот и продуктов их деструкции. Образование специфической окраски. Одним из основных биохимических процессов при посоле мяса для изготовления колбасных и ветчинных изделий являются процессы, связанные с образованием и стабилизацией окраски мяса, обусловленные превращением его пигмента — белка миоглобина (Mb). При посоле мяса миоглобин или оксимиоглобин (МЬ02) в присутствии поваренной соли окисляется и переходит в метмиоглобин (Met. Mb), который придает мясу коричнево-бурую окраску. После варки соленое мясо окрашено в серовато-коричневый цвет, так как в результате распада миоглобина, оксимиоглобина и метмио-глобина при тепловой денатурации образуется коричневый пигмент — гемохромоген. Чтобы окраска соленого сырого и вареного мяса была розовато-красной, к рассолу или в посолочную смесь добавляют нитраты и нитриты. В мясе они подвергаются следующим последовательным превращениям: Восстановительные условия Кислая среда NaNOa---------------------------------- * HONO Восстановительные условия
HONO
NO + Mb При наличии редуцирующих условий нитраты (NaN03 и КИОз) восстанавливаются до нитритов, В слабокислой среде (рН 6, 5—5, 5), характерной для мяса, нитриты под действием ферментов тканевых и микроорганизмов (денитрифицирующих), восстанавливаются с образованием окиси азота. Более кислая реакция среды (рН ниже 5, 5) способствует слишком быстрому распаду нитритов и потере окислов азота в результате улетучивания. Окислы азота, возникшие вследствие распада нитритов, связываются с железом гема в молекуле миоглоби-на или гемоглобина, образуя NO-миоглобин (нитрозомиоглобин) или NO-гемоглобин. Нитрозомиоглобин придает мясу розово-красную окраску. Красный цвет сохраняется и у вареного мяса, так как в результате тепловой денатурации нитрозомиоглобин превращается в денатуриро- Нитрозогемвхрвмвген ч- Иенияурирввдншй гм/и//
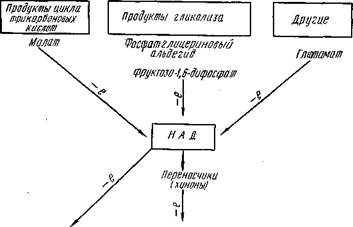
ярко-красный ванный глобин и NO-гемохромоген— пигмент также розово-красного цвета. Оптимальная среда для образования NOMb при рН 5, 6. Превращение миоглобина в оксимиоглобин, метмио-глобин и нитрозомиоглобин сопровождается изменением спектра поглощения раствора этих белков. Процесс образования NO-миоглобина при посоле протекает значительно сложнее, с образованием промежуточных продуктов (рис. 35). В условиях слабокислой среды нитрит непосредственно взаимодействует с оксимио-глобином и в присутствии кислорода воздуха возникает не NO-миоглобин, а метмиоглобин NO" + 2МЬ02--------- > NO; + Mb202 + Va 02 Нитрит Нитрат Метмиоглобин Кроме того, при экспозиции NO-миоглобина в присутствии кислорода возможна следующая реакция: 02+NOMb-----------.-MetMb + NOj,, т. е. образование метмиоглобина. Эта реакция ускоряется в присутствии света и при низком рН. Поэтому при измельчении соленое мясо приобретает коричнево-серый цвет. В отсутствии кислорода (в глубине мяса) нитрит реагирует с Mb и порождает эквивалентное количество метмиоглобина и NO-миоглобина. Показано, что метмиоглобин в растворе в присутствии N0 в анаэробных условиях может переходить в новое своеобразное соединение нитрозометмиоглобин с характерным спектром поглощения. Таким образом, в результате указанных реакций возникает значительное количество метмиоглобина, что мешает образованию NO-миоглобина и поэтому процесс восстановления метмиоглобина при посоле мяса имеет существенное значение. Это возможно в результате взаимодействия протеида с некоторыми соединениями при участии тканевых ферментов. В качестве возможного субстрата-донатора электронов были проверены промежуточные продукты гликолиза, трикарбонового цикла и др. Эффективная роль, как было обнаружено, принадлежит при этом фосфату глицеринового альдегида и фруктозо-6-фос-фату. Восстановление метмиоглобина происходит при помощи НАД. Важное значение в транспорте электронов
принадлежит также диафоразе, хинонам (рис. 36), дегид-рогеназе янтарной кислоты и др.
МетмиоглоВин \ Миоглойин Рис. 36. Схема возможного пути переноса электронов и роль отдельных соединений в восстановлении метмиоглобина в ткани. ного восстановления этой системой метмиоглобина в ни-трозомиоглобин. В модельных опытах на мясе было найдено, что при посоле происходит одновременное снижение содержания SH-групп и нитрита в результате их взаимодействия. Эти же опыты с низкомолекулярными тиолами (цистеин, глютатион) показали, что указанные соединения, реагируя с нитритом, дают нитрозотиолы с характерным спектром поглощения. По-видимому, подобные продукты возникают и в мясе, но влияния их на развитие окраски нельзя считать существенными. Метмиоглобин частично восстанавливается в NO-мио-глобин во время последующего постепенного нагрева (обжарка, варка) соленого мяса, колбасного фарша. Образование NO-миоглобина в этом случае является результатом суммарного действия редуцирующих веществ и каталитического действия особого фермента нитритредуктазы, ускоряющего превращение нитритов в окись азота. Применяя нитриты для посола (нитрит — ядовитый продукт), исходят из минимального количества его, которое необходимо для создания нормальной окраски продукта. Эмпирическим путем работами ВНИИМПа установлено, что эта минимальная доза составляет 5 мг на 100 г мяса. Расход нитрита при посоле зависит не только от количества миоглобина (при этом N0 расходуется не только на прямое образование NO-миоглобина, но и на побочные реакции — образование метмиоглобина и др.), но и от реакции среды, окислительно-восстановительного потенциала, активности ферментов мяса, обсемененности и состава микрофлоры и т. п. Например, чем ниже рН ткани и окислительно-восстановительный потенциал, тем больше расходуется нитрит. Разнообразные превращения нитритов в мясе при посоле в конечном итоге сопровождаются значительным их разрушением, поэтому при изготовлении колбасных изделий свободного нитрата остается 45—25% по отношению к введенному количеству его. Для образования N0 — Mb используется 6—9% N0 (от суммы введенного нитрита); в виде остаточного нитрита в саркоплазме удерживается 21—27% NO, a 4— 8% N0 оказывается тесно связанным с актомиозином. Для создания восстановительных условий в целях лучшего прокрашивания мяса, более полного использования N0 применяют сахара (0, 3—0, 5%). Сами сахара, даже редуцирующие (глюкоза, мальтоза), не создают достаточных восстановительных условий, однако продукты промежуточного анаэробного распада их, образующиеся под действием ферментов бактерий, обладают значительным редуцирующим действием. Кроме того, такая ферментация сахара способствует поддержанию оптимального значения рН (в результате образования молочной кислоты) и окислительно-восстановительного потенциала.
1/г 11—454 Некоторые исследователи считают, что сахара и продукты их распада защищают NO-миоглобин и NO-гемохромоген от окисления. Для посола используют сахарозу или глюкозу, но последняя быстрее вовлекается в окислительные превращения, поэтому применяется только при кратковременном посоле. В случае продолжительного посола применяют сахарозу. В присутствии сахара и нитрита развитие микроорганизмов принимает иногда нежелательное своеобразное направление. При этом наряду с окислами азота из нитритов образуются NH3 и гидроксиламин - о _о + н2о Азотистая Азотноватистая Гидроксиламив кислота кислота
Гидроксиламин (обнаружен в небольшом количестве в рассолах) влияет на дальнейшее направление биохимических превращений в тканях мяса, например он ингиби- Рис. 37. Кривые поглощения, иллюстрирующие процесс превращения нитрозогемоглобина в метгемоглобин при окислении: / — нитрозогемоглобин перед окислением (рН 7, 2); 2 — нитрозогемоглобин после окисления; 3 — метгемоглобин (рН 7, 2). рует каталазу. Гидроксиламин характеризуется большей реакционной активностью и легко вступает во взаимодействие с рядом веществ, в частности с карбонильными соединениями (образуются при окислении Сахаров, жиров), давая новые продукты типа оснований. Нитрозомиоглобин, находящийся в соленом сыром мясе, или нитрозогемохромоген, содержащийся в соленом вареном мясе, т. е. в готовом изделии, при хранении в присутствии кислорода воздуха претерпевает изменения, сопровождающиеся нежелательным частичным или полным обесцвечиванием продукта. Обесцвечивание ускоряется при повышении температуры (15—20°С), под действием света и микрофлоры. Нитрозомиоглобин при этом переходит в метмиоглобин. На Модельном опыте показано превращение NO-гемоглобина в метмиоглобин (рис. 37). Нитрозогемохромоген в чистых растворах также весьма нестоек, но в мясе он устойчивее NO-миоглобина. При хранении готовых изделий все же возможно частичное обесцвечивание продукта в результате окисления NO-гемохромогена. При хранении сваренных соленых мясных изделий иногда появляется зеленая окраска, обусловленная образованием сульфмиоглобина в результате взаимодействия миоглобина с сероводородом или окислительными превращениями миоглобина, вызванными разрывом порфиринового кольца. Для улучшения и сохранения окраски готовых продуктов при посоле применяют некоторые антиокислители, например аскорбиновую кислоту (обычно применяется ее натриевая соль — аскорбинат натрия), которая является сильным восстановителем. Аскорбинат легко взаимодействует с кислородом воздуха, поэтому введенная в ткань кислота в значительной мере защищает пигменты от окисления, поглощая кислород. При взаимодействии аскорбиновой кислоты (при высокой концентрации) и оксимиоглобина наблюдается дегидрирование аскорбината с переходом протонов на протеид и образование нестабильной гидроперекиси миоглобина, которая может затем распадаться на холеглобин и метмиоглобин + Аскорбиновая кислота пМЬОа------------------------------ ► Н2ОаМЬ + Дегидроаскорбиновая / I кислота я/2 MetMb | \ п/2 Холеглобин Метмиоглобин, взаимодействуя с аскорбиновой кислотой, восстанавливается в миоглобин, который в присутствии кислорода превращается в оксимиоглобин
Vs. И* + Аскорбиновая кислота MetMb----------------------------- » Mb -+- Дегидроаскорбиновая кислота |о, МЬ02 Нитрит натрия, взаимодействуя с аскорбиновой кислотой, восстанавливается до окиси азота QH806 + 2HONO---------- » 2NO + 2НаО + С6Н6Ов. Поэтому при посоле мяса с применением аскорбиновой кислоты ускоряется образование NO-миоглобина. Как уже отмечалось, при посоле возможно образование метмиоглобина. Однако с применением аскорбината создаются восстановительные условия, способствующие прямому образованию NO-миоглобина. Это показано спектральными кривыми, полученными на модельных опытах с гемоглобином (рис. 38). Применение аскорбиновой кислоты при нитритном посоле ускоряет процесс цве-тообразования и стабилизирует окраску в процессе хранения готовых изделий.
Рис. 38. Действие аскорбиновой кислоты на раствор оксигемогло' бина: /■ —оксигемоглобшн; 1 — оксигемоглобин +0, 02% NaNOi+0, 1% аскорбиновой кислоты (типичный спектр нитрозогемоглобива); 3 — смесь оксигемогло- бина+0, 02% NaN02 (спектр метмиоглобина); 4 — оксигемоглобин +• 0, 1% аскорбиновой кислоты (раствор зеленовато-коричневого цвета), смесь мет-гемоглобина и холеглобина. Окислительные и гидролитические изменения жиров. При сухом, а отчасти и при мокром посоле мяса, особенно свиного, в присутствии кислорода воздуха частично развиваются процессы автокисления жира, в основном в поверхностных слоях. Благодаря наличию высокоактивных катализаторов главным образом биохимического происхождения (цитохромов, гемоглобина, NO-миоглобина, NO-гемохромогена), а также в присутствии солей, нитратов, нитритов и микрофлоры в этот процесс вовлекаются ненасыщенные жирные кислоты. В результате окисления жира образуются перекиси и карбонильные соединения. При посоле мяса в производстве колбас обнаружено увеличение ТБЧ (см. с. 165). Резко возрастало ТБЧ в момент образования окрашенных продуктов (нитрозо-пигмента), что связано, по-видимому, с появлением летучих соединений, реагирующих с 2-ТБК. Величина ТБЧ может зависеть от присутствия нитрита в системе. Добавление последнего вызывает в начале снижение величины ТБЧ, а затем при посоле она может возрастать. Имеющаяся в тканях липаза не только не ингибирует-ся компонентами посола, но, наоборот, активируется ионами хлооистого натрия и в зависимости от температурных условий может заметно катализировать гидролиз жиров и выделение свободных жирных кислот. Количество перекисей и кислот в соленом мясе обычно увеличивается во время последующего копчения или созревания при комнатной температуре. Изменение вкуса или аромата. В результате разнообразных, еще недостаточно изученных биохимических процессов, протекающих при автолизе соленого мяса, а также вследствие бактериальной деятельности органолепти-ческие свойства мяса изменяются, что сопровождается появлением вкуса и аромата соленого мяса. Особенно заметен специфический вкус и аромат при длительном посоле свиного мяса, получивший название «ветчин-ность». Ветчинный запах появляется после варки соленого свиного мяса. Вещества, являющиеся источником этого запаха, извлекаются при водной экстракции соленой свинины. Максимальный аромат свойственен фракции, высаливающейся сернокислым аммонием в концентрации 50—60%. В создание вкуса и аромата при посоле свиного
10—454 мяса вовлекаются аминокислоты, переходящие в рассол (гистидин, глютаминовая кислота, аланин, серии, таурин, валин, метионин), а также NO-миоглобин. Эти соединения в результате биохимических превращений образуют компоненты, участвующие в формировании вкуса и аромата продукта. К числу веществ, участвующих в создании аромата и вкуса ветчинности, относятся летучие соединения (содержатся в малых количествах). Они могут быть продуктами окислительной деструкции жиров, углеводов и других веществ (аминокислот и азотистых экстрактивных веществ). Относятся они к классам летучих жирных кислот, карбонильных или серусодержащих соединений, аминов и др. Так, при ветчинном посоле было обнаружено накопление летучих оснований и альдегидов. Установлено накопление в беконных рассолах (мокрый посол) ацетил-метилкарбинола (ацетоин) (СН3СОСНОНСН3), который образуется в результате ферментации Сахаров. При окислении ацетилметилкарбинола — 2Н СН3СОСНОНСН3--------- ► СН3СОСОСН8 получается диацетил (СН3СО — СО—СН3) — соединение, обусловливающее натуральный запах < сливочного масла, который выявляется и в других естественных продуктах, например иногда при посоле говяжьего мяса. При посоле окороков обнаружено изменение в соотношении ряда карбонильных соединений — уменьшение высокомолекулярной фракции и нарастание содержания низкомолекулярных соединений. В преобладающем количестве выявлено присутствие ацетальдегида, ацетона, пропионового альдегида. Увеличивается и содержание летучих кислот по мере нарастания срока посола окороков. В создании аромата и вкуса соленого мяса принимают участие тканевые ферменты и, по-видимому, ферменты микроорганизмов. Так, образование ацетилметилкарбинола связано с деятельностью Bacillus subtilis, Bacterium Halobicus, Micrococcus lipoliticus и др. В последнее время выделены чистые бактериальные культуры, которые при посоле вводят в мясо для улучшения вкуса и аромата готового посоленного продукта. Изучение динамики развития отдельных форм микроорганизмов (лактобактерии, микрококки, педиакокки и др.) и динамики свободных аминокислот в течение 46-суточного созревания сырокопченых колбас показало интенсивное накопление аминокислот в первые 6 суток и меньший рост их содержания в последующий период. Сопоставление интенсивности развития микрофлоры и увеличения содержания аминокислот дает основание утверждать, что основная роль в накоплении последних принадлежит катепсинам ткани. Применение Сахаров при посоле способствует получению более вкусного и нежного продукта. Это объясняется тем, что сахара смягчают вкус соли и в то же время являются основным продуктом для развития специфической микрофлоры. В результате действия микрофлоры образуется ряд продуктов промежуточного и конечного окислительного распада гексоз, которые, возможно, участвуют в создании особого вкуса и аромата соленого мяса. Увеличение количества вводимого сахара (свыше 2%) может вызвать нежелательное развитие микрофлоры, что приведет к накоплению избыточного количества кислот и порче продукта (закисание). Отмеченный ранее факт накопления аминокислот при длительном посоле окороков, а также присутствие Сахаров имеет большое значение в образовании летучих компонентов, возникающих в продукте при нагревании в результате активации реакции Майара, о чем будет сказано ниже. Биологическая оценка процесса посола. Происходящая в процессе посола небольшая деструкция части белков с выделением свободных аминокислот и последующим окислением последних обусловливает формирование вкуса и аромата мяса. Хотя подобная деструкция и приводит к незначительной утрате содержания части аминокислот, ее нельзя рассматривать как отрицательное явление. То же относится и к большому снижению содержания некоторых ненасыщенных жирных кислот из фракции липидов. Вместе с тем происходящее при посоле мяса снижение содержания некоторых водорастворимых витаминов — тиамина до 28%, фолиевой кислоты до 35% (потери рибофлавина незначительны, а ниацин сохраняется полностью, так как находится в структуре ткани в связан-
10* ном состоянии) : — нельзя расценивать как положительный фактор. Все изложенное, не вызывая отрицательной оценки процесса посола, свидетельствует о необходимости изыскания путей для снижения или компенсации отмеченных потерь. Иное значение имеет вопрос о роли отдельных ингредиентов, применяемых в процессе посола. В последние годы большое внимание уделяется изучению взаимодействия нитритов и нитратов с другими компонентами тканей, а также реакции организма (реципиента) в целом на поступление этих соединений с колбасными изделиями в качестве пищевых продуктов. При существовавшей до сих пор норме нитрита 10 мг на 100 г мяса при посоле в готовых изделиях было обнаружено содержание нитрита от 1 до 21 мг%. Интересно отметить, что наибольшее содержание нитрита было найдено в свиных сардельках (до 18 мг%) ив зельце красном (до 21 мг%). Увеличение содержания нитрита в готовом продукте по сравнению с исходным сырьем объясняется, по-видимому, тем, что для подготовки отдельных видов сырья использовалась селитра, которая могла в процессе изготовления послужить источником накопления нитрита. При интенсивном кормлении подопытных животных колбасными изделиями было обнаружено, как следствие влияния нитритов, некоторое уменьшение усвоения липи-дов и белков, торможение активности панкреатической липазы, щелочной фосфатазы, энтерокиназы. В крови таких животных увеличивалось содержание метгемогло-бина, что несколько снижало дыхательные функции ее. Если сам по себе нитрат в этом отношении не опасен, то, поступая в кишечник, он с помощью микрофлоры восстанавливается до нитрита и, следовательно, может быть причиной токсических явлений. В колбасных изделиях обнаружен и гидроксиламин, обладающий также способностью стимуляции метгемоглобинообразования. Хотя образование небольшого количества метгемогло-бина под влиянием нитритов не оказывает вредного влияния на здоровый организм, тем не менее отмеченное явление нежелательно. В результате сложных превращений нитритов в мясе установлена возможность появления нитрозаминов. Некоторые из них, например, диметил- и диэтилнитрозами- ны являются не безвредными для организма. В этом же Популярное:
|
Последнее изменение этой страницы: 2016-06-05; Просмотров: 836; Нарушение авторского права страницы

 Установлено, что в тканях восстановление нитрита в окись азота протекает при действии митохондрий, т. е. в этом случае нитрит вместо кислорода замыкает дыхательную цепь. Показана также возможность непосредствен-
Установлено, что в тканях восстановление нитрита в окись азота протекает при действии митохондрий, т. е. в этом случае нитрит вместо кислорода замыкает дыхательную цепь. Показана также возможность непосредствен-
