
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Клеточные и неклеточные формы организации живого: эукариоты, прокариоты, вирусы
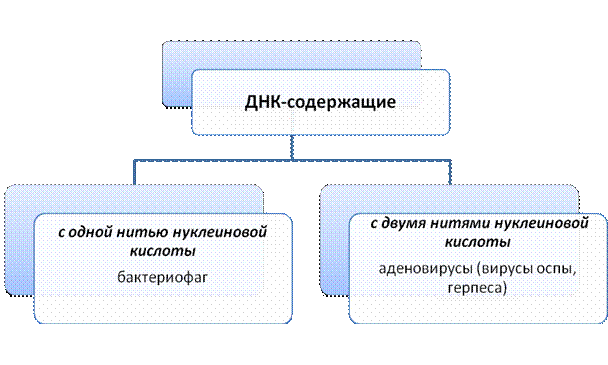
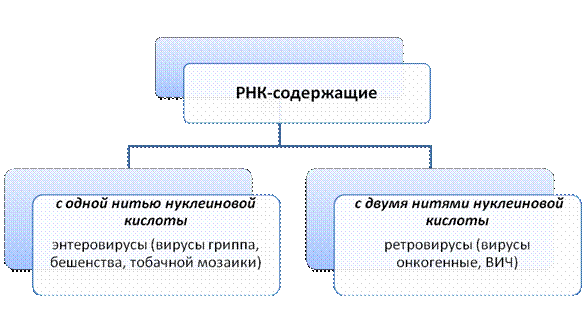
Цитогенетика – это раздел генетики являющийся основополагающим для понимания закономерностей наследственности и изменчивости и требует вспомнить знания биологических предметов (ботаники, зоологии и др.). Цитогенетика – это раздел генетики, изучающий закономерности наследственности и изменчивости на клеточном и молекулярном уровне. Индивидуальное развитие от одной клетки до многоклеточного организма с различными специализированными тканями и органами – это результат последовательного, избирательного включения в активное состояние разных генных участков хромосом в различных клетках. Таким образом, любая клетка многоклеточного организма тотипотентна, то есть обладает одинаковым полным фондом генетического материала, всеми возможными потенциями для проявления его свойств. Но вследствие дифференцировки как результата избирательной (дифференцированной) активности разных генов в клетках по мере развития многоклеточного организма, одни и те же гены в разных клетках могут находиться либо в активном, либо в репрессивном состоянии. То есть в процессе онтогенеза происходит специализация клеток и тканей. При этом у животных организмов такая специализация часто необратима, а у растений даже из отдельных клеток можно получить нормальные растения (вегетативное размножение). У живых организмов существует два типа организации клеток: прокариотическая (доядерная), такая, как у бактерий и сине-зеленых водорослей, которые обычно делятся бинарным образом, то есть простой перегородкой без участия специальных аппаратов деления; и эукариотическая (собственно ядерная), у которых клеточное ядро отделено от цитоплазмы ядерной оболочкой, и нормальным полноценным способом деления является митоз, при котором происходит образование специального аппарата клеточного деления – веретена. Благодаря веретену деления равномерно и точно по двум дочерним клеткам распределяются после деления центромеры 2 хроматиды одной хромосомы, и, таким образом, сохраняется постоянство числа хромосом (2n) и идентичность генетического материала. Неклеточной формой являются вирусы, которые состоят из капсида – защитной белковой оболочки и генетического материала. В качестве наследственного материала вирусы могут содержать 2 вида нуклеиновых кислот и поэтому вирусы подразделяются на ДНК-содержащие и РНК-содержащие.
2. Нуклеиновые кислоты. Структурная модель ДНК ДНК – дезоксирибонуклеиновая кислота – биологическая макромолекула, носитель генетической информации во всех эукариотических и прокариотических клетках и во многих вирусах. В 1928 г. Ф.Гриффит обнаружил у пневмококков явление трансформации (преобразование свойств бактерий). Он показал, что клетки невирулентных штаммов бактерий (шероховатые без капсул) приобретают свойства вирулентных (гладких с капсулами) штаммов, убитых нагреванием. Природа трансформирующего агента была установлена Эвери, Мак-Леодом и Мак-Карти в 1944 г., им оказалась ДНК. Так открытие и изучение трансформации доказало роль ДНК как материального носителя наследственной информации. Трехмерная модель пространственного строения двухцепочечной ДНК была описана в апрельском журнале Nature в 1953 г. Дж.Уотсоном, Френсисом Криком и Морисом Уилкинсом. Эти исследования легли в основу молекулярной биологии, изучающей основные свойства и проявления жизни на молекулярном уровне. Структура ДНК – полимер, структурной единицей которого является нуклеотид. Нуклеотид состоит из азотистого основания пуринового: аденин (А) или гуанин (Г) или пиримидинового: цитозин (Ц) или тимин (Т), углевода дезоксирибозы (пятиугольное сахарное кольцо) и остатка фосфорной кислоты (НРО3). Фосфатные группировки находятся снаружи спиралей, а основания – внутри и расположены с интервалом 34 нм. Двойная спираль ДНК правосторонняя. 10 пар оснований составляют полный оборот 360 градусов, следовательно, каждая пара оснований повернута на 36 градусов вокруг спирали относительно следующей пары. Цепи удерживаются вместе водородными связями между основаниями и закручены одна вокруг другой и вокруг общей оси. В разработке модели ДНК важную роль сыграли наблюдения Чаргаффа (1949) о том, что количественные отношения гаунина всегда равны содержанию цитозина, а содержание аденина соответствует содержанию Тимина. Это положение было названо «правило Чаргаффа»: А=Т; Г=Ц или А+Г/Ц+Т=1, т.е. пропорция пуриновых и пиримидиновых оснований всегда равная. Чаргаффом для характеристики нуклеотидного состава ДНК был предложен коэффициент специфичности, учитывающий долю гуанин-цитозиновых пар: Г+Ц/А+Т или (Г+Ц/А+Т+Г+Ц) Х 100%. Нуклеотиды соединены в полинуклеотидную цепь связями между 5` положения одного пентозного конца и 3` положения следующего пентозного кольца через фосфатную группу с образованием фосфодиэфирных мостиков, т.е. сахарно-фосфатный остов ДНК состоит из 5`- 3` связей. Генетическая информация записана в последовательности нуклеотидов в направлении от 5` конца к 3` концу – такая нить называется смысловой ДНК, здесь расположены гены. Вторая нить направления 3`-5` считается антисмысловой, но является необходимым «эталоном» хранения генетической информации. Антисмысловая нить играет большую роль в процессах репликации и репарации (восстановление структуры поврежденной ДНК). Основания в антипараллельных нитях образуют за счет водородных связей коплементарные пары: А+Т; Г+Ц. Таким образом, структура одной нити определяет последовательность нуклеотидов другой нити. Следовательно, последовательности оснований в нитях ДНК всегда антипараллельны и комплементарны. Принцип комплементарности универсален для процессов репликации и транскрипции. В настоящее время описаны несколько модификаций молекулы ДНК. Полиморфизм ДНК – это способность молекулы принимать различные конфигурации. В настоящее время описано 6 форм, часть которых может существовать только in vitro (в пробирке): В-форма – имеет стандартную структуру, практически соответствующую модели ДНК, которая была предложена Уотсоном, Криком и Уилкинсом, в физиологических условиях (низкая концентрация солей, высокая степень гидратации) является доминирующим структурным типом. А-форма – обнаружена в более обезвоженных средах и при более высоком содержании ионов калия и натрия. Интересна с биологической точки зрения, т.к. ее информация близка к структуре двухцепочечных ДНК, или для ДНК-РНК дуплексов. С-форма – имеет меньше форм оснований на виток, чем В-форма. В этих трех формах могут находиться все ДНК независимо от нуклеотидной последовательности. Следующие формы характерны только для молекул ДНК с определенными последовательностями в парах оснований. D и Е-форма – возможны крайние варианты одной и той же формы, имеют наименьшее число пар оснований на виток. Обнаружены только в молекулах ДНК, не содержащих гуанина. Z-форма – это зигзагообразная форма, с чередованием лево- и правоспиральности. Эта форма выявляется при наличии ряда факторов: высокая концентрация солей и наличие специфических катионов; высокое содержание отрицательных супервитков в молекуле ДНК и других Z-ДНК встречается на участках, обогащенных парами Г-Ц. Показано, что Z-форма ДНК может участвовать в регуляции экспрессии генов как близко расположенных, так и существенно удаленных от Z-участков, а также играть существенную роль в процессах рекомбинации. Шотландский ученый Арнотт предположил: «Было бы удивительно, если бы в живой природе никак не использовалась эта способность ДНК – менять свою форму». Некоторые из форм могут при определенных условиях, связанных с изменениями концентрации солей и степени гидратации, переходить друг в друга, например, А – В; а также Z – В. Предполагают, что взаимные переходы А- и В-форм регулируют работу генов. Показательно, что в ДНК человека имеются участки, потенциально способные переходить в Z-форму. Предполагается, что в клетках человека существуют условия, стабилизирующие Z-форму (Марри и др., 1993). Знание структуры и функции ДНК необходимо для понимания сути некоторых генетических процессов, которые являются матричными. Было ясно, что сама ДНК не может играть роль матрицы при синтезе белков из аминокислот, т.к. почти вся она находится в хромосомах, расположенных в ядре, в то время как большинство, если не все, клеточные белки синтезируются в цитоплазме. Таким образом, генетическая информация, заключенная в ДНК, должна передаваться какой-то промежуточной молекуле, которая транспортировалась бы в цитоплазму и участвовала в синтезе полипептидных цепей. Предположение о том, что такой промежуточной молекулой может быть РНК, стало всерьез рассматриваться сразу, как только была открыта структура двойной спирали ДНК. Во-первых, клетки, синтезирующие большое количество белка, содержали много РНК. Во-вторых, еще более важным казалось то, что сахарофосфатные «скелеты» ДНК и РНК чрезвычайно сходны ( строение РНК ) и было бы легко представить себе, как происходит синтез одиночных цепей РНК на одноцепочечной ДНК с образованием нестабильных гибридных молекул, одна цепь которых представлена ДНК, а другая РНК. Взаимоотношения ДНК, РНК и белка в 1953г. были представлены в виде следующей схемы: Транскрипция трансляция
На последних этапах, предшествующих делению клетки, ядерный материал (хроматин) претерпевает определенные физико-химические изменения, приводящие к конденсации нитеобразных структур ядра. Эти образования немецкий морфолог В.Вальдейер (1888) предложил назвать хромосомами, поскольку они интенсивно окрашивались некоторыми красителями. Хромосомы – это нуклеопротеиновые тела, в которых хранится, передается потомству и реализуется наследственная информация. По иронии судьбы сначала были открыты ядерные структуры, которые в течение многих последующих лет никто не считал хромосомами. В 1881 году Э. Бальбиани описал в клетках слюнных желез хирономуса поперечно-исчерченные ленты. Их назвали " структурами Бальбиани". Только в 1912 году чешский ученый Ф. Рамбоусек предположил, что это специализированные хромосомы. А окончательно это название утвердилось в 1930-1935 гг. (D. Kostoff, T. Painter, H. Midler). Хромосомы, как " окрашивающиеся тела" были открыты в митотически и мейотически делящихся клетках классиками цитологии Флеммингом и Страсбургером (W. Flemming, 1882; Е. Strasburger, 1884). Для каждого вида растений и животных характерны свое число и свои морфологические особенности хромосомного набора, т. е. определенный кариотип. В норме все хромосомы клеток организма эукариот парны, т. е. каждая хромосома имеет своего аналогичного по размеру, форме и особенностям расположения генов гомолога, и составляют двойной (2n), или диплоидный, набор. Только зрелые половые клетки содержат одинарный (1n), или гаплоидный, набор хромосом (от греч. haplous — одинарный). Точное число и структуру отдельных хромосом можно оценить в делящихся клетках на стадии метафазы, когда хромосомы наиболее утолщены и укорочены. В настоящее время наиболее известны три типа хромосом: а.У прокариот в нуклеоиде и в клеточных органеллах у эукариот. б.Хромосомы из делящихся клеток эукариот. в.Интерфазные хромосомы эукариот. Литература 1. Айала, Ф. Современная генетика / Ф. Айала, Дж. Кайгер. – М.: Мир, 1987. – Т.1. – 295 с; Т.2. – 368 с; Т.3. 2. Алиханян, С. И. Общая генетика / С. И. Алиханян, А. П. Акифьев, Популярное:
|
Последнее изменение этой страницы: 2017-03-08; Просмотров: 867; Нарушение авторского права страницы

 репликация ДНК ---------------------- РНК --------------------- белок, где
репликация ДНК ---------------------- РНК --------------------- белок, где