|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
КЛАСС ГОРМОГОНИЕВЫЕ — НОКМОООМОРНУСЕАЕСтр 1 из 29Следующая ⇒
Нитчатые синезеленые водоросли, у которых протопласты соседних клеток сообщаются посредством плазмодесм. Размножаются гормого- ниями, у многих известны споры. Из входящих сюда порядков рассмот Порядок осциллаториевые — 08сШа1опа1е$
Порядок объединяет нитчатые гомоцитные формы. Представителем может служить род осциллатория (ОзсИШопа). Его виды часто образуют сине-зеленые пленки, покрывающие влажную землю, подводные предметы, или плавают в виде толстых кожистых лепешек на поверхности стоячих водоемов. Длинные нити осциллатории сложены из цилиндрических клеток, совершенно одинаковых, за исключением верхушечных, которые по форме могут несколько отличаться от остальных
5. тахьта, содержат много протеинов (60—68 % от сухой массы) и с давних времен используются в пищу населением некоторых районов Африки. В последние годы в ряде стран, в том числе и в СССР, стали заниматься вопросами массового культивирования видов спирулины. - У рода лингбия (1^уп@Ьуа) нити такого же строения, что и у осцилла- тории, окружены хорошо заметным мощным чехлом или влагалищем (рис.38). Порядок ностоковые — N081003168 Порядок объединяет гормогониевые водоросли с гетероцитными нераз* ветвленными нитями или нитями с так называемым ложным ветвлением, Представителями служат следующие роды. частью по гетероцистам. Гормогонии растут только за счет поперечных делений клеток. Большинство видов имеет споры, обычно резко отличающиеся по форме и размерам от вегетативных клеток (рис. 39). У ряда видов (Л. суИпёгьса, А. огугае, А. ьагшЬШз и пр.) доказана способность к фиксации атмосферного азота. У близкого к анабене рода афанизоменон (АрНатготепоп) нити обычно собраны в пучки или чешуйки, видимые невооруженным глазом. Развиваясь в массе в планктоне, вызывает «цветение» воды. Как и мик- роцистис, представляет собой важный продуцент органического вещества в водоемах. В прудовом хозяйстве развитие афанизоменона искусственно стимулируют добавлением в воду калийных и фосфорных удобрений. Род носток (ЫозТос) характеризуется слизистыми или студенистыми колониями разных ра-змеров и формы: от микроскопически мелких до крупных, достигающих величины сливы; от сферических или эллипсоидальных до неправильно распростертых и нитевидных. Слизь разной консистенции, содержит массу извитых нитей или беспорядочно перепутанных, или расходящихся более или менее радиально из центра колонии (рис. 40). Нити похожи на нити анабены: наряду с вегетативными клетками содержат интеркалярные гетероцисты. Размножение—8 посредством гормогониев, которые возникают в результате распада нитей по гетероцистам (при этом гетероцисты отмирают и выпадают) на совершенно прямые участки. Образование гормогониев сопровождается также внутриклеточными перестройками, которые внешне выражаются в пгом, что в большинстве случаев изменяются размеры и форма клеток. Очень часто у бентосных форм, в вегетативном состоянии лишенных газовых вакуолей, эти вакуоли появляются в клетках гормогониев. Гормогонии приобретают подвижность и покидают материнскую колонию, слизь которой к этому времени расплывается (рис. 41, А—Г), а если колония покрыта плотным наружным слоем — перидермом, то этот слой разрывается (см. рис. 43, 2). Гормогонии обычно служат и для распространения вида, так как обладают скользящим движением, и благодаря развитию в клетках газовых вакуолей — лучшей плавучестью. После некоторого периода движения первоначально прямые гормогонии останавливаются, теряют газовые вакуоли (у бентосных видов) и прорастают в спирально извитые нити (рис. 41, Д). При этом клетки гор- могония делятся или косыми (рис. 41, Д; 42, Г), или продольными перегородками (рис. 42, Б). Во втором случае сначала возникает двухрядная нить, а затем при- разъединении определенных клеток этой нити формируется уже зигзагообразная нить, свойственная ностокам |рис. 42, В). Обычно конечные клетки молодой нити превращаются в гетероцисты, выделяется обильная слизь (рис. 41, Е) и таким образом возникает молода» колония, в которой нити дальше растут только за счет поперечных делений клеток. Помимо образования гормогониев у многих ностоков развиваются споры, обычно мало отличающиеся по форме и размерам от вегетативных клеток. Сферические ностоки нередко размножаются почкованием колоний (рис. 43, 5, 6). Самый крупный сферический носток '— сливовидный (Д'ох/ос ргип^огте), в огромных количествах развивающийся в наших северных водоемах, а также
г
Рис. 43. Схема размножения сферического ностока посредством вторичных (образующихся из вегетативных ^ нитей) гормогониев (слева) и почкованием колоний (справа): 1 — зрелая колония, 2 — выход вторичных гормогониев из разорвавшегося перидерма материнской колонии, 3 — проростки гормогониев, 4 — молодая колония, получившаяся из одного проростка, 5 — почкующаяся колония, 6 — две колонии, возникшие в результате почкования
Рис. 44. СаШНпх. А—общий вид нити, Б, В — ложное ветвление: одиночное (Б) и двойное (В)
видны отдельно лежащие гетероцисты (х200); В и Г-—«косы» черных от газовых вакуолей гормогониев, выползающих из колонии на поверхность агара; Д—извитые в результате косых делений клеток гормогонии (Х200); Е — образовавшиеся из гормогониев молодые колонии после выделения слизи (Х100)
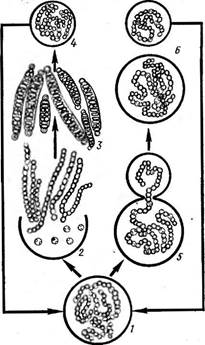
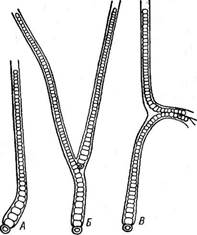
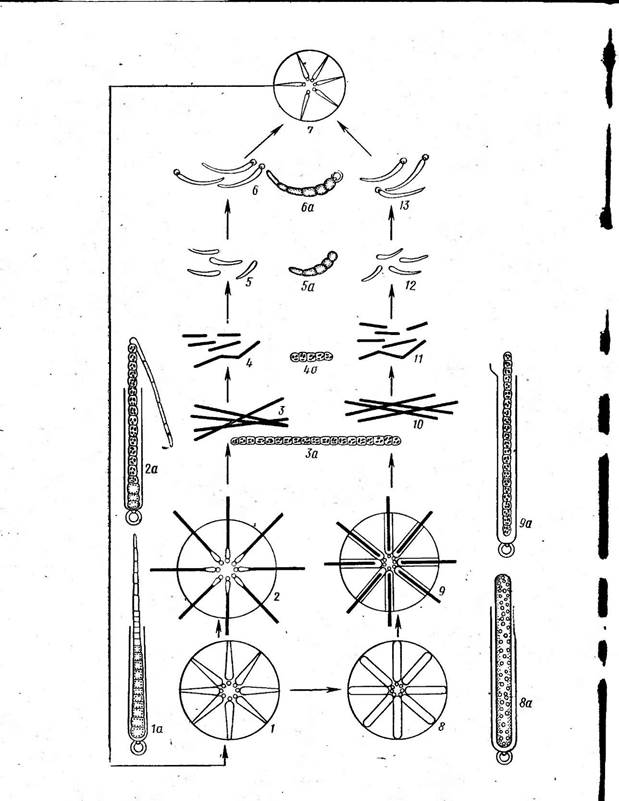
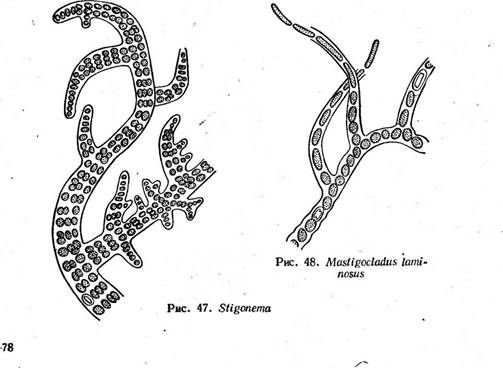
носток с колониями в " виде слизистых тяжей, в которых нити расположены более или менее параллельными рядами (/V. (1аце1Щогте), съедобны. Последний вид, растущий в пустынях и высокогорных областях, употребляется как лакомство. Ностоки способны фиксировать атмосферный азот. У ряда других ностоковых нити асимметричны. Например, у рода калотрикс (СаШНпх) нити бичевидные: на расширенном конце дифференцируется базальная гетероциста (рис. 44, А), противоположный конец утончается и часто заканчивается многоклеточным волоском из отмерших клеток. Нить одета чехлом и может ветвиться. Ветвление ложное и заключается в том, что на протяжении нити в результате отмирания промежуточной клетки происходит разрыв (рис. 44, Б, В). Расположенный ниже разрыва участок, прорывая чехол, продолжает расти вбок, образуя ветвь. Конечная клетка верхнего фрагмента может превратиться в базальную гетероцисту (рис. 44, Б) или же базальный конец верхнего фрагмента может дать вторую боковую ложную ветвь (рис. 44, В). В первом случае ложное ветвление одиночное, во втором — двойное. У родов ривулярия 1апа) и глеотрихия (С1оео1псШа) талломы, как и у ностока, студенистые. Нити асимметричные, как у калотрик- са, окружены общей слизью и расходятся из центра по радиусам. К центру колонии обращены расширенные концы с базальной гетеро- цистой, к периферии — волоски (рис. 45). Размножение калотрикса, ри- вулярии и глеотрихии — посредством гормогониев: при этом " волоски сбрасываются и в образовании гормогония участвуют нижерасположенные клетки, за исключением нескольких базальных. Как и у ностоков, формирование гормогониев сопряжено с изменениями размеров и формы клеток, часто появляются газовые вакуоли. Гормогонии прямые и симметричные, отделяются от основания нити в результате отмирания промежуточной клетки и после некоторого периода движения останавливаются, прорастая в асимметричные нити. При этом на одном конце их клетка превращается в базальную гетероцисту, а на противоположном конце клетки вытягиваются в волосок. У колониальных форм, например у глеотрихии, гормогонии, как правило, собраны в пучки помногу, и молодая колония формируется за счет прорастания многих гормогониев: вокруг развивающихся в асимметричные нити гормогониев выделяется обильная слизь, в которой молодые нити располагаются по радиусам. У ривулярии споры отсутствуют. У некоторых видов калотрикса и у всех видов глеотрихии имеются споры. У глеотрихии длинная спора с зернистым содержимым образуется за счет слияния нескольких клеток, расположенных над базальной гетероцистой. К концу вегетационного периода все нити в колонии образуют споры, вегетативные клетки отмирают и в слизи остаются только споры, у шаровидных видов (например, у О. р1$ит) расходящиеся из центра колонии по радиусам (рис. 46). При помещении таких колоний в благоприятные условия споры более или менее одновременно прорастают. Содержимое их делится с образованием длинных симметричных многоклеточных нитей — с п о р о го р м о г о н и е в или первичных гормогониев, в клетках которых обычно появляются обильные газовые вакуол^ (рис. 46, 9, 9а). Стенка на верхушке споры разрывается, и спорогор- могонии, плотность которых меньше, чем у воды (из-за газовых вакуолей), всплывают на поверхность воды обычно соединенные пучками, где и претерпевают дальнейшее развитие, подобное развитию гормогониев (вторичных), образующихся из взрослых вегетативных колоний. Схематически цикл развития О. рьзит представлен на рис. 46. Порядок стигонемовые — 5Н§опета(а1е$ Для представителей характерны гетероцитные нити с настоящим ветвлением. Примерами могут служить роды стнгонема (8Н§опета) и мастигокладус {МазИ& ос1айиз). Нити стигонемы обладают верхушеч-
ным ростом. На некотором расстоянии от верхушки ветвей клетки претерпевают продольные деления, и нить становится многорядной (рис. 47). Мастигокладус с единственным видом М. 1аттози8 имеет сложный ветвящийся гетероцитный таллом. Представляет собой типичную термальную водоросль, распространенную по всему свету (рис. 48). * * * г Особенность индивидуального" развития гормогониевых водорослей заключается в том, что в ходе развития из гормогониев взрослых особей они проходят ряд неодинаковых морфологических стадий, или статусов, сходных со зрелыми особями других родов. Так, все гормо- гониевые водоросли в состоянии гормогониев отвечают признакам рода осциллатория (симметричная гомоцитная нить). При дальнейшем развитии разные роды различаются по статусам, и число последних тем- больше, чем сложнее организация таллома данного рода, чем больше имеется разных морфологических признаков, формирующихся независимо друг от друга. Так, если у рода.осциллатория весь жизненный' цикл сведен к одному состоянию (осциллаториеподобному), то у рода лингбия к статусу осциллаториеподобному прибавляется еще статус лингбиеподобный, для которого характерен толстый чехол вокруг гомо- цитной нити. У рода калотрикс наблюдаются состояния оециллаторие- подобное (гормогонии) и калотриксоподобное (зрелые особи с асимметричными гетероцитными нитями). У рода ривулярия в жизненном цикле уже три состояния: осциллаториеподобное, калотриксоподобное и ривуляриеподобное, а у рода глеотрихия — четыре: осциллаториеподобное, калотриксоподобное, ривуляриеподобное и глеотрихиеподоб- ное — взрослый таллом со спорами (табл. 3).
Наконец, у маетигокладуса, имеющего сложный гетероцитный ^таллом с дифференцировкой вегетативных клеток, различающихся по форме в разных частях таллома с разными типами ветвления (истинного, ложного), удается установить в жизненном цикле уже десять статусов. На разных морфологических стадиях (или статусах) гормогониевые - водоросли могут задерживать свое развитие, длительно существовать в таком состоянии и даже размножаться посредством гормогониев. Относительно связей синезеленых водорослей с другими отделами низших растений говорилось выше. По строению клетки и химизму клеточной оболочки синезеленые водоросли, несомненно, близки к бактериям. Содержание же в клетках хлорофилла «а» сближает рх с эука- риотными водорослями, в первую очередь с красными, имеющими сходные с синезелеными дополнительные пигменты, локализованные в форме фикобилисом на поверхности одиночно расположенных тила» кондов. И в той и в другой группе.отсутствуют жгутиковые стадии. Однако если обе эти группы и имеют общих предков, то очень отдаленных, о которых трудно составить какое-нибудь. представление. Синезеленые водоросли — очень древние, гораздо более древние, чем красные, их находки известны из докембрийских отложений. К этему периоду относятся уже находки как безгетероцистных форм, так и форм с гетероцистами. Начиная с палеогена в пресноводных отложениях синезеленые водоросли представлены уже современными родами. Таким образом, синезеленые водоросли, достигнув высокой степени дифференциации еще в далекие геологические времена, с тех пор эволюционировали сравнительно мало и дошли до наших дней, не претерпев заметных морфологических изменений. По-видимому, первичные синезеленые водоросли были представле-» ны одноклеточными, не одетыми оболочками формами, от которых в самостоятельных направлениях эволюционировали хамесифоновые и хроококковые. Между этими группами прямых родственных связей нет. От хроококковых в результате неполного (незавершенного) деле-
I I Первичные гетеро'цистные водоросли Первичные гормогониевые водоросли (без гегероцисг)- ♦ I Исходные одноклеточные формы Рис. 49. Схематическое изображение вероятных путей эволюции основных групп гормогониевых водорослей • ' ния клеток могли возникнуть первичные осцилляториеподобные гомо- цитные формы, от которых эволюция пошла по трем основным путям: один из них привел к становлению современных осцилляториевых водорослей, два других, направленных в сторону разделения функций между клетками первичного нитчатого таллома, — к современным ностоко- вым и стигонемовым водорослям. Схематически эволюция основных групп гормогониевых водорослей представлена на рис. 49. Приведенная '(кажущаяся нам наиболее вероятной) концепция эволюции гормогониевых водорослей — не единственная, имеются и другие взгляды; в частности, некоторые авторы выводят гормогониевые ■ водоросли от ха- месифоновых. ■ * * * Распространены синезеленые водоросли повсеместно, растут там, где не могут произрастать никакие другие растения и первыми заселяют вновь образующиеся поверхности Земли, например вулканические острова. Большинство синезеленых обитают в пресных водах, меньше —в море. В планктоне прудов, озер, медленно текущих рек виды родов микроцистис, анабена, афанизоменон, глеотрихия часто вызывают «цветение» воды. Планктонные виды обычно имеют в клетках газовые вакуоли, благодаря чему всплывают на поверхность воды. ТпсНоАезтшт егуНггаеит (от избытка фикоэритрина красного цвета) вызывает «цветение» воды в Красном море, что и послужило причиной для названия этого моря. Многие синезеленые водоросли развиваются вне воды, например на почве полупустынь юго-востока СССР (главным образом ностоки), на скалах во влажных тропиках (глеокапса, сцитонема). Среди них широко распространен симбиоз: представители ряда родов представляют собой гонидии (фикобионты) лишайников (носток, стигонёма, сцитонема, калотрикс), живут в корнях высших растений, являются внутриклеточными симбионтами некоторых жгутиковых, корненожек и др. Синезеленые водоросли имеют большое значение в жизни человека, как положительное (азотфиксация, съедобные), так и отрицательное (токсичные, возбудители «цветения» воды, которые при отмирании портят воду и могут вызвать массовое отмирание — замор рыбного населения водоема). |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 265; Нарушение авторского права страницы
 Рис. 34. Ражкеппета (ЕпЛопета). Эндоспоро- образование
Рис. 34. Ражкеппета (ЕпЛопета). Эндоспоро- образование
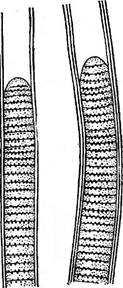 Рис. 38. 1упЁЬуа. Общий вид нити
Рис. 38. 1упЁЬуа. Общий вид нити
 Рис. 37. 8р(гиИпа. Общий вид инти
Рис. 37. 8р(гиИпа. Общий вид инти
 Рис. 36. ОхИШопа. Общий вид нити
Рис. 36. ОхИШопа. Общий вид нити
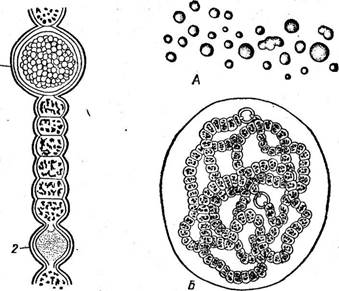 Рис. 39. АпаЬаепа. Общин вид нити:
1 — спора, 2 — гетероциста
Рис. 39. АпаЬаепа. Общин вид нити:
1 — спора, 2 — гетероциста
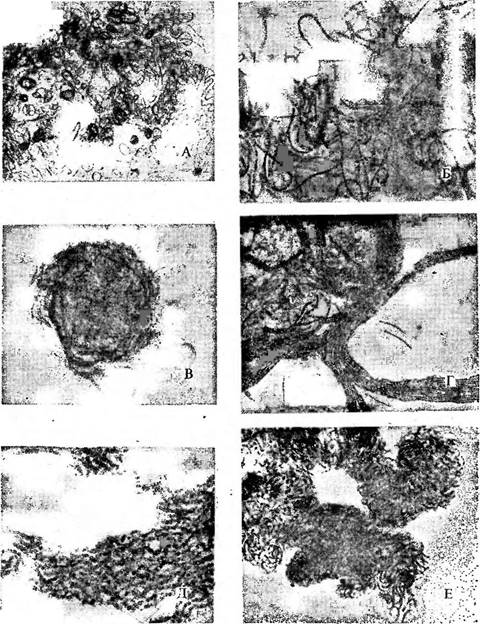 Рис. 41. Размножение ностока посредством гормогониев. Ыо& 1ос рипсН/огте. А — в расплывшейся слизи колонии видны разматывающиеся нити, клетки которых еще до распада на гормогонии содержат газовые вакуоли (Х62); Б—нити с клетками, заполненными газовыми вакуолями, распадаются по. гетероцистам на гормогонии, в слизи еще
Рис. 41. Размножение ностока посредством гормогониев. Ыо& 1ос рипсН/огте. А — в расплывшейся слизи колонии видны разматывающиеся нити, клетки которых еще до распада на гормогонии содержат газовые вакуоли (Х62); Б—нити с клетками, заполненными газовыми вакуолями, распадаются по. гетероцистам на гормогонии, в слизи еще
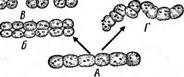 Рис. 42. Развитие из гормогония (А) извитых нитей ностока (В, Г) в результате продольных (Б), или косых (/') делений клеток
Рис. 42. Развитие из гормогония (А) извитых нитей ностока (В, Г) в результате продольных (Б), или косых (/') делений клеток