
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Порядок немалионовые — №таНопа1е8
Для представителей характерно отсутствие ауксилярных клеток, гонимобласты развиваются непосредственно из оплодотворенного карпогона или из его дочерней клетки. Виды порядка жцвут главным обра-
а а. Б Рис. 55. ВЫгасНоврегтит. А — внешний вид таллома; Б — часть таллома с цистокарпиями зом в морях, но встречаются и в пресных водах, например роды лема* неа (Ьетапеа) и батрахоспермум (Ва1гасНо5регтит). Род леманеа (Ьетапеа) встречается в быстро текущих речках с холодной водой. Таллом, имеющий вид неветвящейся щетинки 10—15 см длиной и 1 мм толщиной, темно-фиолетового или оливково- бурого цвета- с узловатыми вздутиями. Прикрепляется к субстрату при помощи подошвы из стелющихся нитей. По оси таллома проходит одна нить из вытянутых бесцветных клеток. От верхней части каждой клетки центральной оси отходит мутовка большей частью из четырех расходящихся по радиусам ветвей. Базальные клетки каждой такой ветви крупные, вытянутые. От их дистального (морфологически верхнего) конца отходят ветви второго порядка, которые в свою очередь многократно ветвятся. Конечные разветвления срастаются в многослойную кору. Ее наружные клетки мелкие и заполнены хроматофорами, внутренние—крупные и почти бесцветные. От проксимальных (морфологически нижних) концов основных клеток радиальных ветвей отходят коровые нити, окутывающие центральную нить (рис. 54, А, Б, В). Антеридии образуются группами из поверхностных клеток коры (рис. 54, Г). Карпогониальные ветви, несущие карпогоны, развиваются из внутренних клеток коры. Брюшко карпогона находится около внутренней поверхности коры, а трихо- гина прорывает кору и высовывается наружу (рис. 54, 5). После оплодотворения из брюшной части карпогона формируются пучки го- нимобластов, врастающие в полость таллома. Карпоспорангии образуются целыми цепочками, карпоспоры скапливаются внутри полости таллома и освобождаются после разрушения коры таллома. Согласно последним исследованиям Мань (1967), при развитии из зиготы (оплодотворенного брюшка карпогона) гони- мобластов мейоз не наблюдается и, таким образом, карпоспорофит и карпоспоры — диплоидны. Карпоспоры прорастают с образованием диплоидного нитчатого, состоящего приблизительно из 20 клеток предростка, в верхушечной клетке которого происходит редукционное деление. Из апикаль- - ной клетки после мейоза развивается гаплоидное растение — гаметофит леманеи, на котором образуются половые органы. Так что базальная часть таллома леманеа диплоидна.
Род батрахоспермум (Ва1гаско5регтит, рис. 55, А) встречается в реках с чистой прозрачной водой, в озерах. Таллом в виде сильно разветвленного, слизистого на ощупь кустика оливково-зеленого или стального цвета (избыток фикоцианина). Ось неограниченно нарастает за счет деятельности верхушечной клетки и состоит из одного ряда вытянутых бесцветных клеток. От каждой из них непосредственно под поперечной перегородкой возникает мутовка ветвей ограниченного роста, которые сложены из мелких, богатых хроматофорами клеток. Конечные клетки ветвей могут вытягиваться в длинные волоски. Из базальных клеток боковых ветвей (ассимиляторов) развиваются ветви неограниченного роста. Кроме того, базальные клетки мутовок ветвей дают начало кортикальным (коровым) нитям, которые растут вдоль клеток главной оси, совершенно скрывая их, и у некоторых видов, переплетаясь, образуют многослойную обвертку. Клетки коро- вых нитей дают начало вторичным мутовкам боковых ветвей, которые вставляются между уже имеющимися. На боковых ветвях ограниченного роста (ассимиляторах) формируются половые органы. После оплодотворения из брюшка карпогона вырастают ветвящиеся гонимобласты, .конечные клетки которых дают карпоспоры, собранные в тесную группу— цистокарпий (рис. 55, Б). Из карпоспор развиваются ползучие Рис. 57. ЫетаИоп. А — внешний вид таллома; Б — поперечный разрез таллома нити, от которых вертикально отходят разветвленные нити совеем иного внешнего вида, чем таллом батрахоспермума. Эта стадия была описана под родовым названием шантранзия (СНап1гап51а). Она может размножаться посредством моноспор (рис. 56). При благоприятных условиях из верхушечных клеток С/гап^гапзш-предростка могут вырастать типичные талломы батрахоспермума. Согласно исследованиям Мань, СкаМгапзьа-стацию батрахоспермума, так же как и базальную часть таллома леманеа, следует рассматривать как диплоидное образование, соответствующее тетраспорофиту других красных водорослей. Талломы леманеа и батрахоспермума построены по одноосевому типу. Примером красной водоросли с многоосевым строением может служить морской род немалион (А1етаНоп). В слабоветвящихся розового цвета слизистых талломах немалиона центральная часть занята целым пучком продольных нитей, состоящих из вытянутых бесцветных клеток, от которых радиально расходятся обильно ветвящиеся, богатые хроматофорами нити — ассимиляторы, соединенные слизью мягкой консистенции (рис. 57). 5
Половые органы развиваются, как и у батрахоспермума, на ассимиляторах. Карпоспорофит и карпоспоры у немалиона, как у леманеа и батрахослермума, диплоидны. У одного из видов ЫетаНоп юег- т(си1аге) наблюдалась гетероморфная смена поколений: макроскопического гаметофнта и микроскопического нитевидного спорофита (тетраспорофита). Порядок криптонемиевые — Сгур1опегтпа1е& В отличие ют предыдущего порядка здесь имеются ауксилярные клетки, которые развиваются до оплодотв< зрения карпогона и рассеяны по таллому на известном расстоянии от карпогона. Из карпогона после оплодотворения к ауксилярным клеткам растут более или менее длинные многоклеточные соединительные, или ообластемные, нити: - После слияния ауксилярной клетки с клеткой ообластемной нити и перехода диплоидного ядра клетки нити в ауксилярную клетку возникают гонимобласты (рис.58). Развивающиеся на гонимобластах (карпоспорофите) карпоспоры содержат диплоидное ядро и прорастают в диплоидные растения — тетраснорофиты, производящие только органы бесполого размножения — тетраспорангии. При образовании тетраспор происходит мейоз и гаплоидные тетраспоры прорастают в гаплоидные растения — гаметофиты, несущие половые органы.. Гаметофит и тетраспоро- фит внешне (морфологически), не различаются. Таким образом, у крип- тонемиевых, как и у большинства красных водорослей, имеется изоморфная смена генераций, усложненная диплоидным карпоспоро- фитом. ■ Представитель порядка — род дюреиеа (Оийгезпауа), распространенный в южных морях. Таллом имеет вид разветвленных слизистых кустиков розового цвета. Анатомически он очень похож на батрахо- спермум, так как у него вдоль таллома проходит одна центральная ось из крупных бесцветных клеток, которые несут мутовки сильновет- вящихся коротких боковых нитей из клеток, богатых хроматофорами. Конечные разветвления боковых нитей, переплетаясь и срастаясь, образуют рыхлую кору. На женских гаметофнтах от основания боковых ветвей ограниченного роста вырастают карпогониальные ветви с карпого- нами; такое же положение занимают и ветви, некоторые клетки которых превращаются в ауксилярные. На мужских гаметофитах на конечных веточках развиваются антеридии. На тетраспорофитах из конечных клеток боковых ветвей формируются тетраспорангии. После мейоза содержимое их делится поперечными перегородками на четыре тетраспоры, расположенные друг над другом.
Среди криптонемиевых, как и среди немалионовых, встречаются формы не только одноосевого, но и многоосевого строения. Примером может служить род кораллииа (СогаШпа), распространенный во всех морях, особенно в тропиках. Вертикально стоящие кус- Рис. 59. СогаШпа. А — внешний вид; Б— членики с промежуточными сочленениями; В — концептакул с антеридиями; Г•—концептакул с тетраснорангиямн тики ветвятся преимущественно-в одной плоскости (рис. 59, А). Ветви состоят из сильно пропитанных известью члеников, соединенных друг с другом сочленениями с малым содержанием извести!, что придает всему кустику гибкость, помогающую противостоять действию волн. Если декаль- цинировать таллом (подействовать соляной кислотой), то можно видеть, что вдоль ветвей проходит много (целый пучок) нитей, неограниченно нарастающих и сложенных из длинных бесцветных клеток. От этих осевых нитей по радиусам отходят ветви, срастающиеся в кору. В местах сочленений (рис. 59, Б) центральные оси остаются неразветвленными. На верхушке пучок центральных нитей разделяется обычно на три части, которые далее растут, как ветви. Органы размножения у кораллины (антеридии, карпогоны, тетраспоры) развиваются всегда на разных индивидах (мужском, женском гаметофитах, на тетраспорофитах) на концах ветвей в особых углублениях — концептакулах, сообщающихся с внешней средой посредством узкого отверстия (рис. 59, В, Г). Пропитанные известью талломы коралл лины наряду с такими же талломами сифоновых водорослей участвуют в образовании коралловых рифов.
У рода литотамнион (ЬИНоОгаттоп, рис. 66) таллом, также пропитанный известью, образует на камнях плотные розоватые корочки. Особенно распространен в северных морях, часто на больших глубинах.
ственной близости от последнего, т. е. имеется прокарпий (рис. 61, Б). Длинных ообластемных нитей здесь не образуется: брюшко карпогона или непосредственно сливается с рядом расположенной ауксилярной клеткой (клетками), или через короткие выросты, формируя цисто- карпии (рис. 61, В). Простейшие представители, например род кал- литамнион (СаНЫНаттоп), имеют таллом в виде разветвленного кустика, сложенного из одного ряда многоядерных клеток (рис. 61, Л). Другие представители имеют внешне более сложные, морфологически расчлененные талломы, которые, однако, легко свести к одноосевому типу. Например, у рода делессерия (Ое1е8$епа) таллом в виде ярко- красных листьев с перистым жилкованием (рис. 62) образован срастанием боковых ветвей одной главной оси. В качестве типичного представителя порядка подробнее можно рассмотреть род полисифония (Ро1у81ркота), широко распространенный как в южных, так и в северных морях. Таллом имеет вид разветвленного темно-малинового кустика, ветви которого увенчаны верхушечной клеткой, отчленяющей сегменты. На некотором расстоянии от верхушечной клетки сегменты претерпевают последовательные деления продольными перегородками, в результате чего от центральной клетки отчленяется ряд периферических перицентральных клеток. Таким образом, в более старых частях таллома клетки центральной оси (центрального сифона) окружены перицентральными коровыми клетками (рис. 63, А, Б). У некоторых видов перицентральные клетки делятся с образованием многослойной коры.
На гаметофитах (мужском и женском) половые органы возникают на особых веточках таллома, так называемых трихобластах, имеющих моносифоновое строение — из одного ряда клеток, не окруженных перицентральными клетками. Плодущая. часть мужского трихобласта становится полисифоновой, т. е. клетки отчленяют продольными перегородками перицентральные клетки, которые становятся материнскими клетками антеридиев. От них отпочковываются мелкие антери- диальные клетки (рис. 63, Г; 64, 4—7). Клетки плодущей части женского трихобласта отчленяют пять перицентральных клеток, от одной из которых развивается карпогониальная ветвь с карпогоном, а после оплодотворения отчленяется ауксилярная клетка; остальные перицентральные клетки дают начало нитям, которые образуют вокруг про- карпия и развивающихся карпоспор (цистокарпия) псевдопаренхиматозную оболочку с отверстием на вершине (рис. 63, Д; 64, 15). Из цистокарпия по созревании выпадают карпоспоры, которые развиваются в тетраспорофиты, морфологически сходные с гаметофитами,
На тетраспорофите образуются тетраспорангии, в которые превращаются перицентральные клетки. В тетраспорангиях развиваются по четыре расположенные по углам тетраэдра тетраспоры (рис. 63, В; 64, 17—19).
Таким образом, у иолисифонии (и других представителей порядка церамиевых), как и у криптоиемиевых, наблюдается изоморфная смена генераций, усложненная диплоидным карпоспорофитом. Красные водоросли представляют собой естественную единую, несмотря на многообразие, и весьма древнюю группу; остатки их извест- ' ны из силура и девона. Как указывалось, по набору пигментов, одиночному расположению тилакоидов, отсутствию жгутиковых стадий из всех эукариотных отделов водорослей красные ближе всего к сине- зеленым, от которых они, однако, резко отличаются строением клетки и наличием полового процесса. Отсутствие жгутиковых стадий у современных красных водорослей не давало возможности искать их предков среди первичных жгутиконосцев. Недавно (I. Зкпоп-ВюЪагй-Вгеаий, 1972) появилось сообщение о существовании в клетках красной водоросли ВоппеШщвопха Нить- $ега крипт — видоизмененных скрытых жгутиков. Этот факт, возможно, позволит по-новому подойти к проблеме филогенетических связей красных водорослей с другими отделами водорослей, ве т тре& уе! проверки и подтверждения Что касается эволюции в пределах самого отдела красных водорослей, то из двух классов: бангиевых и флоридей, первый, несомненно, более примитивный. Здесь у карпогона еше не выработалась типичная форма, и он мало отличается от обычных вегетативных клеток. После оплодотворения содержимое карпогона непосредственно делится на карпоспоры. Флоридеи представляют собой более эволюцнонировав- ' шую группу с карпогоном, снабженным органом улавливания сперма- циев — трихогиной. Из класса флоридей наиболее простой порядок немалионовые, у которых нет ауксилярных клеток, и гонимобласты, на которых развиваются карпоспорангии, образуются непосредственно из брюшной части оплодотворенного карпогона. Криптонемиевые представляют следующий шаг прогрессивной эволюции: они имеют ауксилярные клетки, способствующие увеличению продукции карпоспор, так как образуется не один цистокарпий, а много — пз числу ауксилярных клеток. При разбросанных беспорядочно по таллому ауксилярных клетках необходимы более или менее длинные ообластемные нити. Наивысшей ступени эволюции достигли церамиевые, у которых имеется прокарпий и ауксилярные клетки дифференцируются только после того, как произошло оплодотворение. Непосредственное соседство карпогона и ауксилярной клетки (клеток) в прокарпии облегчает образование цистокарпиев. Этот порядок наиболее богат видами. * * * За исключением немногих, преимущественно простейших представителей (ВШгасНоврегтит, Ьетапеа и др.), живущих в быстро текущих речках с чистой водой, красные водоросли — морские обитатели. Они всегда прикреплены к камням, ракушкам, другим водорослям, являясь, как правило, эпифитами, однако известны и эндофиты, и даже паразиты. Многие красные водоросли живут на больших глубинах, и глубоководные формы отличаются особенно яркой красной окраской. Нередко в пределах одного и того же вида глубоководные особи имеют ярко-малиновые талломы, тогда как мелководные особи окрашены в желтоватый цвет. Этот факт пытается объяснить теория Энгельмана (см. с. 253), согласно которой преобладание той или иной окраски на разных глубинах связано с силой и окраской световых лучей, достигающих разных глубин. Красные водоросли находят довольно большое применение в хозяйстве человека: из них вываривают студенистое вещество — агар- агар, применяемое в кондитерской и ряде других отраслей промышленности, а также в лабораторной -микробиологической технике для приготовления твердых питательных сред. Некоторые, например порфира; употребляются в пищу. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 320; Нарушение авторского права страницы
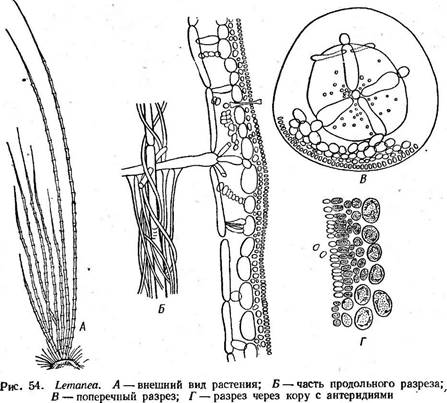
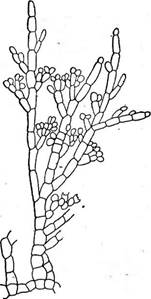 Рис. 56. Стадия СНап- Iгаизш с моноспорами
Рис. 56. Стадия СНап- Iгаизш с моноспорами
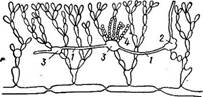 Рис. 58. БитопИа. Схема развития ообластемной нити (1) из карпогона (2) и ее последовательное слияние с аук- силярными клетками (<? ) с образованием гонимобластов (4)
Рис. 58. БитопИа. Схема развития ообластемной нити (1) из карпогона (2) и ее последовательное слияние с аук- силярными клетками (<? ) с образованием гонимобластов (4)
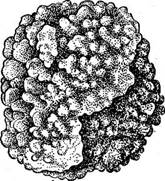 Рис. 60. ШИо{Натмоп. Внешний вид таллома
Рис. 60. ШИо{Натмоп. Внешний вид таллома
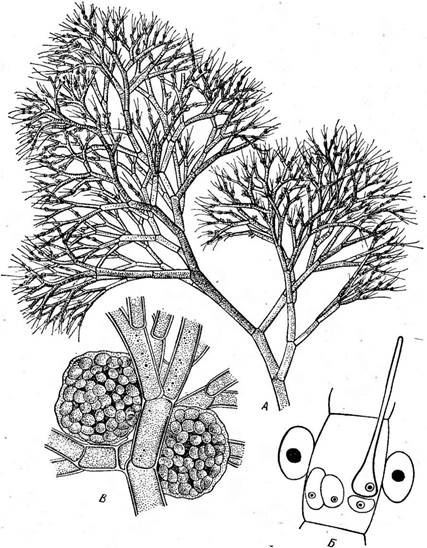 Рис. 61. СаИННшппюп. А — общий вид таллома; Б—прокар- пий; В — цистокарпии
Рис. 61. СаИННшппюп. А — общий вид таллома; Б—прокар- пий; В — цистокарпии
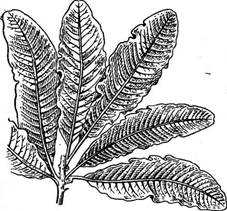 Рис. 62. Ое/евзепа. Внешний вид таллома
Рис. 62. Ое/евзепа. Внешний вид таллома
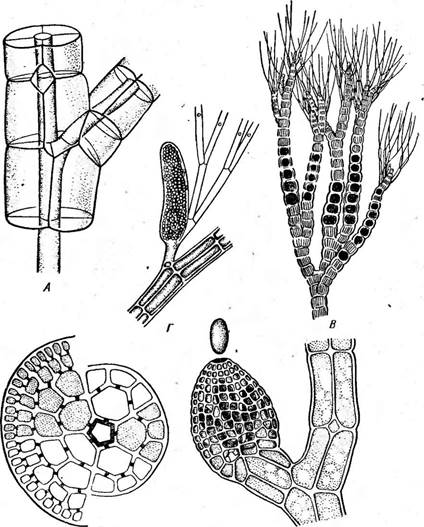 Б Д
Рис. 63. Ро1утркоша. А — схема строения ветви с однослойной корой; Б — поперечный разрез ветви с многослойной *корой; В ■ — тетраспорофит с т етраспорангиями; Г — собрание аитеридиев; Д — зрелый цистокарпий
Б Д
Рис. 63. Ро1утркоша. А — схема строения ветви с однослойной корой; Б — поперечный разрез ветви с многослойной *корой; В ■ — тетраспорофит с т етраспорангиями; Г — собрание аитеридиев; Д — зрелый цистокарпий
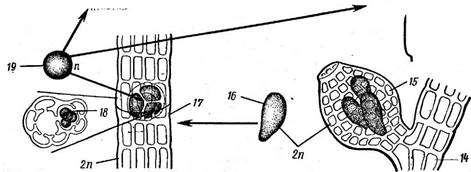 Рис. 64. Ро1уз1рНота. Схема развития:
■ 1 — центральная клетка, 2— перицентральные клетки, 3 — мужской гзметофит, 4—собрание анте- ридиев, 5 — центральная клетка, 6— материнская клетка ашеридиев. 7 — антеридии, 8 - - карпогоин- а льна я ветвь: 9 — карпогон с яйцеклеткой, 10— трихогина, II — спермаций; 12—ауксилярная клетка, отчленяющаяся от поддерживающей клетки карпогониальной ветви после того, как произошло оплодотворение брюшка карпогона, 13 — слияние ауксилярной клетки с брюшком оплодотворенного карпогона, 14—женский галет офит. 15 — цистокарпий, одетый оболочкой, 16 — карпоспор-.* 17 — тетра- спорофнт, 18 — тетраспорангий с тетраспорами, 19 — тетраспора
Рис. 64. Ро1уз1рНота. Схема развития:
■ 1 — центральная клетка, 2— перицентральные клетки, 3 — мужской гзметофит, 4—собрание анте- ридиев, 5 — центральная клетка, 6— материнская клетка ашеридиев. 7 — антеридии, 8 - - карпогоин- а льна я ветвь: 9 — карпогон с яйцеклеткой, 10— трихогина, II — спермаций; 12—ауксилярная клетка, отчленяющаяся от поддерживающей клетки карпогониальной ветви после того, как произошло оплодотворение брюшка карпогона, 13 — слияние ауксилярной клетки с брюшком оплодотворенного карпогона, 14—женский галет офит. 15 — цистокарпий, одетый оболочкой, 16 — карпоспор-.* 17 — тетра- спорофнт, 18 — тетраспорангий с тетраспорами, 19 — тетраспора