
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ОТДЕЛ ЗЕЛЕНЫЕ ВОДОРОСЛИ — СНЬОКОРНУТА
Представители отдела характеризуются травянисто-зеленой окраской хлоропластов, зависящей от преобладания хлорофиллов «а» и «Ь» над каротиноидами: а- и р-каротином, лютеином, неоксантином, вио- лаксантином, зеаксантином, антераксантином. Хлоропласты окружены оболочкой из двух мембран, «хлоропластная эндоплазматическая сеть» отсутствует. Ламеллы состоят из двух — шести или многих слившихся тилакоидов, образующих граны, периферические тилакоиды отсутствуют, генофоры рассеянные. Пиреноид, погруженный в строму хлоропласта, пронизан тилакоидами. Запасной полисахарид крахмал откладывается внутри хлоропласта — вокруг пиреноида и в строме. Глазок располагается внутри хлоропласта и не связан со жгутиковым аппаратом. Жгутики в числе двух, четырех, реже многих одинаковой длины (изоконтные) и одинакового строения (изоморфные), гладкие или покрытые тонкими мастигонемами, у некоторых — чешуйками. Клетка подавляющего большинства зеленых водорослей - поверх цито- плазматической мембраны покрыта целлюлозной оболочкой. Размножение вегетативное, бесполое и половое. Имеются все возможные циклы развития (гаплонты, диплонты, изо- и гетероморфная смены генераций). * Этот отдел занимает среди низших растений особое положение. Окраска зеленых водорослей такая же, как и у высших растений. В качестве запасного вещества в клетках, как и у высших растений, обычно откладывается крахмал. У многих зеленых водорослей наблюдается свойственное высшим растениям правильное чередование поколений: полового и бесполого и, наконец, ряд этих водорослей покинули водную среду и перешли к наземному существованию. Все эти особенности приводят к выводу, что именно среди зеленых водорослей надо искать родоначальников высших растений. Представители отдела большей частью распространены в пресных водах, хотя имеются и морские виды. Во многих системах отдел зеленых водорослей делят на три класса. Класс собственно зеленые водоросли, или равножгутиковые, — СЫо- горЬусеае, 1восоп1ае характеризуется бесполым размножением обычно зооспорами с двумя — четырьмя, реже многими изоконтными и изоморфными жгутиками, обращенными вперед, иногда апланоспорами. Половой процесс гологамныи, изогамный, гетерогамный и оогамный. В этом классе весьма полно представлены ступени морфологической дифференцировки таллома (все, кроме ризоподиальной), которые обычно кладутся Ъ основу деления класса на порядки. Класс конъюгаты — Согци^йорЬусеае характеризуется отсутствием в жизненном цикле жгутиковых стадий, так как бесполое размножение отсутствует, а половой процесс — конъюгация. Класс харовые водоросли — СЬагорЬусеае включает крупные водоросли со сложной морфологической дифференцировкой нитчатого таллома. Бесполого размножения зооспорами или апланоспорами нет. Половой процесс оогамный. Половые органы — оогонии и антеридии — многоклеточные, сложного строения. КЛАСС СОБСТВЕННО ЗЕЛЕНЫЕ, ИЛИ РАВНОЖГУТИКОВЫЕ, ВОДОРОСЛИ — СНЬОРОРНУСЕАЕ, 150С01ЧТАЕ Этот класс занимает центральное положение среди всех зеленых водорослей. В соответствии со ступенями морфологической дифференцировки таллома класс делится на порядки. Порядок вольвоксовые — Уо|уоса! е8 Сюда относятся монадиые, т. е. снабженные жгутиками, одноклеточные, колониальные и ценобиальные зеленые водоросли, подвижные в течение вегетативной жизни. 4 Типичный представитель-^одноклеточная водоросль из рода хламидомонада (СМатуйотопаз, рис. 65), многочисленные виды которого обитают в лужах, канавах и других мелких пресных водоемах. При их массовом развитии вода нередко принимает зеленую окраску. Сферическая или эллипсоидальная клетка, одетая оболочкой, плотно прилегающей к протопласту или (у более старых особей) несколько отстающей от него в задней части, несет на своем переднем конце два жгутика, с помощью которых она плавает в воде. Протопласт содержит одно ядро, обычно чашевидный хроматофор, в который погружены пиреноид и глазок, и сократительные вакуоли, находящиеся в передней
части клетки. Схематическое изображение клетки хламидомонады при наблюдении в электронном микроскопе дано на рис. 22. При подсыхании водоема хламидомонады легко переходят в так называемое пальмеллевидное состояние: при этом клетки теряют жгутики, стенка их ослизняется, и в таком неподвижном состоянии клетки р& змножаются. Стенки дочерних клеток также ослизняются, и таким образом получается система вложенных друг в друга слизистых обверток, в которых группами расположены неподвижные клетки (рис. 65, Б). При этом клетки сохраняют свои пульсирующие вакуоли и глазки. При перенесении в воду клетки снова вырабатывают жгутики и возвращаются к монадному состоянию. Однако легкость перехода из пальмеллевидного состояния в монадное у разных видов неодинакова. Есть хламидомонады (СН. Ыети), у которых большая часть жизни проходит в пальмеллевидном состоянии. Близки к хламидомонаде род картерия (Саг1епа), клетка которого несет четыре жгутика, и род хлорогониум (СН1ого§отит), имеющий сильно вытянутое в длину веретеновидное, иногда игловидное тело с двумя жгутиками. Несколько более по строению клетки отличается от хламидомонады род гематококк (Наета1ососсиз), характеризующийся сильным ослизнением внутренних слоев стенки. " При этом протопласт образует цитоплазменные отростки, тянущиеся через ос- лизнениые слои стенки к наружному, более плотному пограничному слою..Хроматофор чашевидный или сетчато продырявленный, пиреноидов два или несколько. Имеются глазок, пульсирующие вакуоли, ядро и два жгутика (рис. 66, А). Гематококк легко переходит в пальмеллевидное состояние, образуя при этом цисты: клетки округляются, вырабатывают плотно прилегающую стенку, содержимое переполняется астаксантином (гёматохромом), благодаря чему приобретает кирпично- красную окраску. Цисты выдерживают высыхание, а при смачивании прорастают с образованием зооспор, которые выходят наружу через разрыв стенки цисты (рис. 66, Б, 'В). Сначала они окрашены в кир- пично-красяый цвет, но постепенно астаксантин исчезает и с периферии клетки обнаруживается зеленый. хроматофор. Однако красная окраска сохраняется обычно в центре клетки в течение нескольких поколений клеток, размножающихся бесполым путем. Только при длительной культуре, препятствующей образованию цист, удается получить чисто зеленые особи гематококка.
В благоприятных условиях хламидомонада и остальные одноклеточные роды интенсивно размножаются бесполым путем. Обычно клетка при этом останавливается, и про- Рис. 66. НаетаЮсоссш. А-вегета- топласт, несколько отстав от стенки, тивная особь; Б— циста; В — прора- последовательно делится продольно станке цисты вегетативными оссбями на две, четыре или восемь частей
(рис. 65, В). Эти дочерние клетки вырабатывают жгутики и выходят наружу после ослизнения стенок материнской клетки в виде зооспор, одетых оболочкой и отличающихся от материнской клетки только меньшими размерами. После непродолжительного роста они сами приступают к такому же размножению. Имеется и половой процесс. У большинства видов одноклеточных вольвококсовых он изогамный. Гаметы образуются внутри материнской клетки так же, как зооспоры, но в большем количестве (32—64) и соответственно меньших размеров (рис. 67, А, Б). У некоторых одноклеточных вольвоксовых наблюдается гетерогамия и даже оогамия. Так, у СН1атус1отопаз Ьгаипи русский ботаник И. Н. Горожанкин описал гетерогамный половой процесс, при котором крупные женские гаметы, образующиеся в материнской клетке в числе четырех, сливаются с мелкими мужскими гаметами, возникающими в клетке в числе восьми (рис. 67, В). Оогамию также впервые описал И. Н. Горожанкин у СН. сосс^ега. У этого вида одни клетки теряют жгуты и превращаются в неподвижные яйцеклетки, в других клетках формируется большое количество мелких двужгутиковых мужских гамет, которые и оплодотворяют яйцеклетки (рис. 67, Г, Д, Е). Позднее оогамный половой процесс наблюдался еще у нескольких видов одноклеточных вольвоксовых — СН1ого§отит ооцатит, Саг1епа (уеп^аги и некоторых других.
У многих видов хламидомонады наблюдается гетероталлизм, т. е. половой процесс возможен только при смешении «-(-» и «—» гамет и начинается с образования групп — агрегации (скопления) гамет противоположных знаков. Если гаметы одного знака движутся беспорядочно, равномерно заполняя все поле зрения, то при добавлении в такую каплю гамет противоположного знака в поле зрения появляются отдельные скопления гамет — группы. В группах гаметы сначала склеиваются попарно своими жгутиками, после чего активируется автоли- тическая система клеток, растворяющая стенки гамет (гаметы вольвоксовых в отличие от голых гамет большинства водорослей имеют - стенку). Только гаметы, освободившиеся от своих клеточных стенок, сливаются, образуя зиготы. Некоторые ■ ^г? о^ет* вещества; ингибирующие автолиз кле- точных стенок гамет хламидомонады, подавляют и образование зигот, так как половой процесс не происходит. Как показали электронно-микроскопические исследования, у некоторых видов (СМатуйотопаз гетНагйШ) га- Рис. 68. СЫатуйотопаз гетНагМи. меты соединяются посредством опло- Соединение гамет посредством опло- дотворяющей трубки (вырастающей от дотпоряющей трубки (от) переднего конца между жгутиками од ной из сливающихся гамет, рис. 68), у других видов такая трубка отсутствует. Таким образом, агглютинация жгутиков, активация автолитической системы и автолиз клеточных стенок гамет, слияние протопластов представляют собой последовательные этапы в половом процессе хламидомонад.
Вскоре после соединения голых протопластов гамет вокруг них образуется новая клеточная оболочка и их ядра сливаются, формируя диплоидное копуляционное ядро зиготы.
В последние годы с помощью электронного микроскопа удалось проследить дальнейшее поведение пластид после копуляции гамет. Зиготы получались в результате скрещивания дикой исходной формы Ск. геткагйШ, хлоропласты которой плотно заполнены стопками тилакоидов с желтым мутантом, имеющим пластиду, совершенно лишенную тилакоидов. Таким образом, благодаря четким различиям в структуре пластид обеих родительских форм можно- было проследить их дальнейшее поведение после копуляции гамет. После слияния клеток и ядер наблюдалось слияние пластид: в основании пластид в участке, близком к пиреноиду, возникал тонкий мостик, а затем пластиды сливались. Сначала в слившемся хлоропласте зиготы можно различить две морфологически различающиеся части: одна часть с развитыми тилакоидами от хлоропласта дикого типа, другая часть без тилакоидов, принадлежащая пластиде желтогр мутанта. Позднее в хлоропласте зиготы уже не удается различить частей родительских пластид: мем-
\ бранные-структуры — тилакоиды хлоропласта- дикого типа равномерно распределяются по всему хлоропласту зиготы. При прорастании зиготы до мейоза хлоропласт делился на четыре части, и образовавшиеся молодые зооспоры все имели хроматофоры дикого типа. Ряд одноклеточных вольвоксовых в отличие от хламидомонады и близких к ней родов характеризуется отсутствием клеточной стенки. Так, род дуналиелла (ОипаЫеИа, рис. 69), встречающийся в- соленых озерах с высокой концентрацией солей, по форме клетки и внутреннему строению очень похож на хламидомонаду: тот же хроматофор в виде чаши, в утолщенном дне которой находится пиреноид, ярко-красный глазок, ядро, от переднего конца клетки отходят два жгута. Клетки дуналиеллы голые, одетые только цитоплазматической мембраной. Кроме того, у солоноводных видов дуналиеллы в протопласте отсутствуют пульсирующие вакуоли." В соленых озерах часто вместе с дуналиеллой встречается другой род — астеромонас (А8{& готопаз=51еркапор(ега), характеризующийся, как и дуналиелла, голыми клетками обратнояйцевидной или конусовидной формы. Клетка имеет шесть выступов или ребер, проходящих по всей длине тела, поэтому при рассматривании с полюса она выглядит как шестилучевая звезда (рис. 70). От переднего конца клетки отходят два жгутика. Как дуналиелла, так и астеромонас размножаются в основном продольным делением клеток. Известен и половой процесс в виде хологамии, когда копулируют клетки, внешне не отличающиеся от вегетативных. У многих вольвоксовых клетки, имеющие строение хламидомонады или гематококка, соединяются в подвижные ценобии неодинаковой формы, состоящие из различного числа клеток. Из ценобиальных представителей вольвоксовых чаще всего встречаются роды гониум (Оо- пшт), пандорина (Рапйоппа), эвдорина (Еийоппа), вольвокс (Уо1- уох). Гониум образует пластинчатые колонии из клеток, расположенных в один слой. У наиболее широко распространенного вида С отит рес1ога1е (рис. 71, А) колонии состоят из 16 клеток, которые соединяются оттянутыми концами своих стенок, образуя пластинку или таб-
личку, окруженную снаружи бесцветной слизью. Клетки в этом пластинчатом ценобии ориентированы' таким образом, что все передние концы, несущие жгутики, обращены в одну сторону. 'У остальных перечисленных родов ценобиальных вольвоксовых колонии представляют собой слизистые эллипсоиды или сферы, по периферии которых в один слой располагается различное (в зависимости от рода и вида водоросли) число клеток. % У пандорины и эвдорины ценобии микроскопически мелкие, образованы клетками, имеющими строение хламидомонады. Повсеместно встречающийся вид Рапйоппа тогит представляет собой слизистый эллипсоид, по периферии которого под плотным слоем слизи (инволюкрумом) располагаются 16 клеток. Клетки лежат в общем инволюкруме очеНь тесно, давят друг. на друга, вследствие чего имеют граненую форму (рис. 72), и только часть их поверхности, обращенная, кнаружи, выпуклая. Общая полость колонии очень мала.
У эвдорины (рис. 73, А) эллиптические ценобии крупнее, также слизистые, в которых по периферии располагается большее число клеток (32—64), свободно лежащих-в слизи. Центр ценобия занят жидкой слизью. Жгутики, отходящие от передних, направленных к периферии ценобия концов клеток, проходят сквозь одевающую всю колонию плотную слизистую Ьбвертку через находящиеся в ней тонкие канальцы и работой своих свободных частей приводят колонию в движение.
Рис. 74. Уо/солг аигеиа. Дочерние шары внутри материнского Рис. 75. Уо/солг [г[оЬа1ог. Д — строение стенки шара с поверхности (/) и в разрезе (2); Б — колония с яйцеклетками (/) и сперматозоидами (2) Гониум, пандорина, эвдорина, как и одноклеточные представители вольвоксовых, при постепенном подсыхании могут переходить в паль- меллевидное состояние. В таком состоянии эти водоросли размножаются и при культивировании на твердых (агаризированных) питательных средах. При перенесении в жидкие среды клетки вырабатывают жгутики и выходят из слизи в виде зооспор, которые развиваются в соответствующие ценобии.
Наиболее высоко организован вольвокс (рис. 74). Он представляет собой крупный, достигающий 2—3 мм в диаметре шар, одетый тонким инволюкрумом, под которым по периферии шара в один слой располагаются клетки. Число их колеблется от 500 до 60000. Внутренняя полость шара занята жидкой слизью. Отдельные клетки ценобия вольвокса построены по тому же типу, что и клетки гематококка, т. е. внутренние слои стенки с возрастом очень сильно ослизняются, в результате чего протопласт каждой клетки значительно удален от самого наружного, не ослизненного слоя стенки. Более Плотные наружные стенки соседних клеток, соприкасаясь, образуют полигональный узор, иногда хорошо видимый, если смотреть на ценобий с поверхности. Протопласты же соседних клеток.раздвинуты далеко друг от друга. От протопласта, занимающего центр клетки, в одной плоскости, параллельной поверхности всей колонии, по радиусам отходят более или менее многочисленные цитоплазматические отростки, проходящие сквозь ослизненные внутренние слои стенки до самых наружных плотных слоев стенки (рис. 75, Л). Цитоплазматические отростки соседних клеток соответствуют друг другу и связаны между собой плазмодес- мами. У одних видов (Уо& ох §1оЪа1ог) эти отростки толстые, грубые, в них заходят лопасти хроматофора; при рассматривании таких ценобиев с поверхности протопласты кажутся звездчатыми (рис. 75, Л). У других видов (У. аигеиз) цитоплазматические отростки тонкие, едва различимые. В оптическом разрезе клетки или сильно сжаты, конической формы (V. @1оЬа1ог), или более или менее округлых очертаний (V. аигеиз) и на наружной поверхности несут по два жгутика, выходящих наружу через канальцы в инволюкруме. При бесполом размножении ценобиальных вольвоксовых образуются дочерние колонии. При этом протопласт- клеток претерпевает продольные деления, формируя полный комплекс клеток будущего ценобия, после освобождения которого дальнейшие деления клеток прекращаются до наступления новой репродуктивной фазы. Например, бесполое размножение гониума происходит следующим образом: ценобий теряет подвижность, и содержимое каждой клетки делится в продольном направлении последовательно на две, четыре, восемь и, наконец, 16 частей с образованием пластинки; каждая клетка вырабатывает жгутики. Таким образом, внутри стенки материнской клетки формируется новый ценобий, отличающийся от материнского только меньшими размерами. При ослизнении стенки материнских клеток дочерние ценобии освобождаются, и на месте одной 16-клеточной колонии образуются 16 новых ценобиев. Они растут только за счет увеличения размеров клеток, но не их числа. После некоторого периода роста дочерние ценобии снова приступают к бесполому размножению.
При бесполом размножении родов, имеющих сферические ценобии (Рапйоп- ■ па, Еийогига, Уо/уох и др.), содержимое клетки, приступающей к образованию Рис. 77. Уо1г> ох ц1оЬа1ог. Развитие дочернего шара, стрелки указывают полярность клеток (подробное объяснение в тексте) молодой колонии, также несколько отстает от стенки и делится продольными перегородками последовательно на две, четыре, восемь и 16 частей (рис. 73, 5). Стадия 16-клеточной пластинки из-за своего сходства с гониумом получила название тонической. Дальнейшие продольные деления, в результате которых устанавливается определенное для каждого рода и вида число клеток, сопровождаются загибом краев получившейся клеточной пластинки (тонического состояния) до тех пор, пока не образуется полая сфера с маленьким незамыкающимся отверстием, направленным к наружной поверхности родительского ценобия (рис. 76). На этой стадии клетки " ориентированы таким образом, что их передние концы, на которых должны возникнуть жгутики, направлены к центру сферы. Нормальная ориентировка клеток с передними концами, направленными кнаружи, дости-
гается путем полного выворачивания наизнанку незамкнутой сферы, и только после этого ее отверстие замыкается. Клетки образуют жгутики, и получается маленький сферический ценобий (рис. 77). У видов гониума, пандорины и одного вида эвдорины (Е. е1едапз) все клетки, составляющие ценобий, могут образовывать дочерние ценобии при бесполом размножении и гаметы при половом. У остальных видов эвдорины и у рода плевдорина (Р1еийог1па), отличающегося от эвдорины большими размерами сферических колоний, сложенных из 128 клеток, можно проследить постепенную стерилизацию клеток колоний: часть клеток ценобия, обычно передних, в числе четырех, двенадцати, половины или даже больше теряют способность к образованию дочерних колоний (рис. 78). Наиболее резко такая дифференциация клеток колонии на чисто вегетативные и репродуктивные выражена у вольвокса, где из громадного числа клеток, доставляющих колонию, к образованию дочерних колоний способны очень немногие (8—10). Клетки вольвокса, способные давать дочерние колонии, носят специальное название г о н и д и и. Половой процесс у ценобиальных вольвоксовых, кай и у одноклеточных, различается по форме: у гониума и пандорины наблюдается изогамия, у эвдорины — гетерогамия, у вольвокса — оогамия. У гониума в каждой клетке образуется по 16 гамет, которые выходят в воду, где и копулируют. У пандорины гаметы образуются в результате распада молодых колоний на отдельные 'клетки, функционирующие как гаметы. Эвдорина (Еийогта е1е@апв) — форма двудомная, т. е. можно различить колонии женские и колонии мужские. В женских колониях клетки непосредственно превращаются в крупные, снабженные жгутиками гаметы, которые, однако, неподвижны и остаются в слизи женской колонии (ввиду неподвижности женских гамет половой процесс эвдорины часто считают оогамией). В мужских колониях в результате продольных делений клеток возникают пучки вытянутых двужгутиковых сперматозоидов, которые выходят наружу и плавают в воде. Вступив в контакт с женским ценобием, пучки распадаются на отдельные сперматозоиды, которые проникают в слизь женской колонии и оплодотворяют женские клетки (см. рис. 73, В). У вольвокса половой процесс оогамный. Содержимое мужских клеток — антеридиев— делится с образованием пучков двужгутиковых желтоватых сперматозоидов (см. рис. 75, Б). Содержимое женских клеток — оогониев — превращается в одну крупную яйцеклетку. Формирующаяся в результате полового процесса зигота одевается толстой многослойной стенкой и у всех вольвоксовых переходит в состояние покоя. При ее прорастании редукционно делится диплоидное копуля-ционное ядро, образуя четыре гаплоидных ядра и, как правило, четыре зооспоры. Так, у многих видов хламидомонады зигота прорастает с образованием четырех зооспор. У гониума (Сотит рес1ога1е) при прорастании зиготы возникает маленькая четырехклеточная юве- нильная колония (см. рис. 71, Г, Д). Из каждой клетки такой колонии при дальнейшем бесполом размножении формируется уже 16-клеточ- ная колония, типичная для этого вида. В других случаях не все из четырех ядер, образующихся в результате редукционного деления, остаются жизнеспособными, часть их дегенерирует. Например, у пандо- рины, эвдорины при прорастании зиготы остается только одно жизнеспособное ядро и соответственно развивается только одна жизнедеятельная зооспора. То же наблюдается при прорастании зигот у некоторых видов вольвокса (см. рис. 73, Г). Цитологических исследований редукционного деления ядра в прорастающей зиготе сравнительно немного (например, у вольвокса). Гораздо больше косвенных доказательств такого деления получено с помощью генетического анализа зооспор, образующихся при прорастании зиготы. Так, А. Пашеру удалось скрестить два вида хламидомонад, различающихся по строению вегетативных клеток и зигот. Получившиеся в результате скрещивания зиготы имели строение, промежуточное между зиготами родительских видов. При прорастаний таких гибридных зигот в каждой образовалось по четыре зооспоры. Они были изолированы, и при дальнейшем их бесполом размножении были получены четыре культуры, из которых две оказались тождественными с одним из родительских видов, а две другие — с другим. Этот опыт показал, что разъединение признаков гибрида наблюдается при прорастании зиготы, согласно же цитогенетическому учению такое разъединение происходит при редукционном делении диплоидного ядра. Еще больше аналогичных данных получено относительно разделения полов у раздельнополых, или гетероталличных, форм. Экспериментальное изучение полового процесса многих вольвокео- вых показало, что среди них есть обоеполые, или гомоталличные, виды и раздельнополые, или гетероталличные. Пример первых (гомоталлич- ных)—многие виды хламидомонады и вольвокса: у них половой процесс может происходить в культуре, полученной из одной особи, размножившейся бесполым путем. Такие культуры называют клонами. Таким образом, у гомоталличных форм половой процесс возможен в пределах одного клона. У некоторых видов хламидомонады, гониума, пандорины, эвдорины, дуналиеллы половой процесс в пределах одного клона невозможен, так как все особи в пределах клона тождественны в половом отношении. Половой процесс наступает 'только при соединении особей, неодинаковых в половом отношении, из разных клонов. Такие формы называют гетероталличными. Для них в ряде случаев было показано, что разъединение половых свойств происходит в момент прорастания зиготы. Для гониума (Сотшп рес1ога1е) это было продемонстрировано опытами Е. Шрейбера. Он изолировал четырех- клеточные колонии, развивающиеся при прорастании зиготы (рис. 71, Г, Д). В каждой клетке такой колонии через несколько дней формировалась- нормальная 16-клеточная колония. Каждая из четырех первых вегетативных колоний также изолировалась и служила для получения клона. Таким образом Шрейбер выделил четыре клона из четырех клеток, образовавшихся в прорастающей зиготе. Попарное смешение этих клонов позволило установить, что два из них относятся к одному полу, два — к другому. Ввиду изогамии гониума особи противоположного пола обозначаются как «-(-» и «—». В той и другой паре при смешении половая реакция не наступает, копуляция - наблюдается лишь при соединении особей разного пола: «+» и «—Таким образом, у гониума уже" при прорастании зиготы оба пола устанавливаются в равном числе. Разделение." полов в момент прорастания зигот — косвенное доказательство редукционного деления: при этом одни ядра получают потенцию одного пола (+), другие — потенцию противоположного пола (—). При дальнейшем бесполом размножении знак пола не меняется. Определение пола в момент прорастания зиготы, связанное с редукционным делением, называется г е н о т и п и ч ее к и м 1 (или гаплогенетическим). У гомоталличных обоеполых видов при прорастании зиготы тоже имеет место редукционное деление (например, у вольвокса это было доказано цитологически), но оно не влечет за собой разделения полов. Выходящие из зиготы зооспоры потенциально несут оба пола, и лишь в их потомстве наступает половая дифференцировка, которая не* связана с редукционным делением и больше определяется внешними условиями. Такое определение пола называется фенотипи- ческим (или физиологическим). # * * Дуналиелла, ' астеромонас (представители семейства ОипаНеИа- сеае) — обитатели соленых вод. Они приспособились к перенесению любых концентраций солей, однако область концентраций, при которых клетки не только остаются живыми, но и размножаются, -очень мала и резко ограничена для каждого вида. Известны и некоторые виды хламидомонад, которые встречаются в соленых озерах с очень высокой концентрацией солей. Среди колониальных вольвоксовых есть и некоторое количество морских форм (например, ОНтапзьеНа). Однако подавляющее большинство вольвоксовых -— пресноводные организмы. Колониальные вольвокеовые иногда встречаются в мелких лужах вместе с хламидомонадами, йо чаще обитают в более крупных постоянных водоемах с чистой водой без каких-либо значительных органических загрязнений. Многие виды хламидомонад также приурочены к чистым водам, однако ряд видов живет в воде, сильно загрязненной органическими веществами. Таковы СЫатуйотопав еНгепЬег^и, СН. то- пасИпа, СН. гетНаг(1Ш. Экспериментальные исследования этих видов в чистых культурах показали, что лучше всего они растут на средах с добавлением органических веществ: если в среде в качестве источ-- ников углерода присутствуют сахара, то возможен их рост в темноте. Наилучшим источником азота оказались аминокислоты. СН. еНгепЬег^и и некоторые другие виды, отчасти перейдя к сапро- трофному питанию, не утратили способности к фотосинтезу, т. е. питание таких видов миксотрофное (смешанное). У многих других вольвоксовых, которые приобрели способность к питанию органическими веществами, фотосинтез оказался утраченным. Эти сапротрофные формы встречаются в сильно загрязненных органическими веществами водах. При переходе от авто- к гетеротрофному питанию ассимиляционный аппарат этих форм претерпел более или менее полную редук- ■ 1 Генотипическое определение пола было установлено В. Лерхе у БипаИейа & аНпа* 110 а -з Рис. 79. Параллельные ряды окрашенных и бесцветных форм среди вольвоксовых, А. Б — ВипаЧеНа (А — вегетативная особь, Б — продольное деление клетки); В. Г — ЯуаНеПа (В — вегетативная особь, Г — размножение посредством продольного деления клетки); Д, Е — СЫатуЛотопаз (Д— вегетативная особь, Е — зооспорообразование); Ж, 3 — Ро1у1ота (Ж — вегетативная особь, 3 — зооспорообразование); . И — СМогодо- гйит; К — НуаЬдотит цию. Таким образом возникли бесцветные, полностью лишенные хром а- тофоров и хлорофилла формы. Бесцветные формы известны и в других порядках водорослей, но у вольвоксовых особенно наглядно можно продемонстрировать параллельные ряды окрашенных и бесцветных форм (рис. 79). Так, формой, параллельной дуналиелле (рис. 79, А, Б), ■ отличающейся от нее только отсутствием хроматофора, будет род НуаНеИа. Оба эти рода характеризуются голыми двужгутиковыми клетками, размножающимися продольным делением (рис. 79, В, Г). Род Ро1у1ота (рис. 79, Ж, 3) отличается от СМатуйотопаз (рис. 79, Д, Е) отсутствием хлорофилла. Аналогичным образом различаются роды СМогоцотит (рис. 79, И) и Нуа1оцоп1ит (рис. 79, К)- Порядок тетраспоровые — Те1га$рога1ев К вольвоксовым очень близок порядок тетраспоровых (нередко рассматриваемый как семейство Те1ха5рогасеае в порядке вольвоксо- 'вых), характеризующийся пальмеллоидным типом строения таллома. Представителями могут служить роды апиоцис- тис (АрюсузИз) и тетраспора (Те1газрога). Таллом апиоцистиса представлен грушевидными слизистыми колониями, прикрепленными к другим водорослям (рис. 80). У тетраспоры мешковидные, нередко бесформенные, крупные, видимые невооруженным глазом слизистые колонии. В слизи как у апиоцистиса, так и у тетраспоры находятся клетки, по строению весьма напоминающие хламидомонаду. Каждая клетка несет по два неподвижных жгутикоподобных отростка — псевдоцилии. Электронно-микроскопическое исследование показало, что эти структуры— производные жгутиков.. Их базальные тельца и корни почти идентичны с таковыми хламидомонады; переходная зона имеет звездчатый узор, характерный для жгутиков зеленых водорослей. Однако свободная часть органеллы не имеет центральных микротрубочек и В-трубочки периферических дублетов большей частью очень коротки. Таким образом, под мембраной псевдоцилий микротрубочки (микрофибриллы) имеют расположение 9—0 вместо характерного для типичных жгутиков (9—9) +2. Каждая клетка колонии апиоцистиса и тетраспоры может превратиться в зооспору, при этом вместо псевдоцилий вырабатываются два настоящих жгутика. Зооспоры выходят из слизи и после некоторого периода подвижности прорастают, образуя новые колонии. Наблюдается и половой процесс в виде изогамии. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 342; Нарушение авторского права страницы
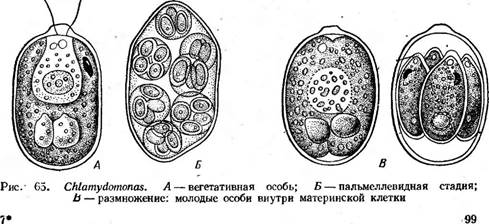
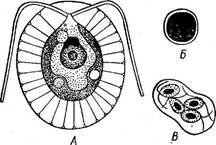
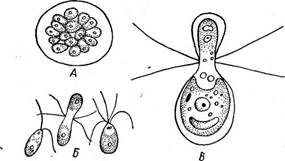 Рис. 67. СМатуйотопаБ. Половой процесс: А— образование изогамет; Б—изогамия у СЛ. зШпп; В — гетерогамия у СЛ. Ьгаипи; Г — оогамия у СЛ. соса[его; Д — мужская гамета СЛ. соса}ега; Е — зигота
Рис. 67. СМатуйотопаБ. Половой процесс: А— образование изогамет; Б—изогамия у СЛ. зШпп; В — гетерогамия у СЛ. Ьгаипи; Г — оогамия у СЛ. соса[его; Д — мужская гамета СЛ. соса}ега; Е — зигота
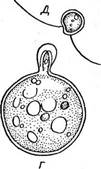

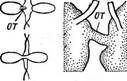
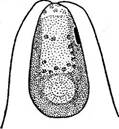 Рис. 69. ОипаИеПа. Вегетативная особь
Рис. 69. ОипаИеПа. Вегетативная особь
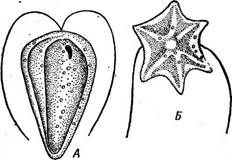 Рис. 70. А81еготопа$, А — клетка сбоку;
Б — клетка с полюса
Рис. 70. А81еготопа$, А — клетка сбоку;
Б — клетка с полюса
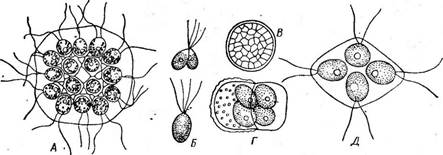 Рис. 71. Сотит рес(ога1е. А — ценобий; Б — копуляция гамет; В — зигота;
Г, Д — прорастание зиготы
Рис. 71. Сотит рес(ога1е. А — ценобий; Б — копуляция гамет; В — зигота;
Г, Д — прорастание зиготы
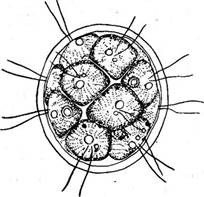
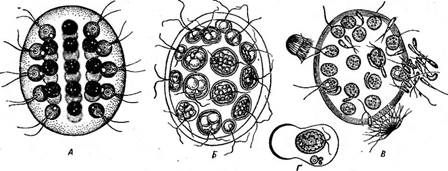 Рис. 73. ЕиЛоппа е1едап& А — ценобий; Б— развитие дочерних '-колоний внутри клеток материнской; В — оплодотворение; Г — прорастание зиготы: одна нормальная зооспора и три абортивные (справа)
Рис. 73. ЕиЛоппа е1едап& А — ценобий; Б— развитие дочерних '-колоний внутри клеток материнской; В — оплодотворение; Г — прорастание зиготы: одна нормальная зооспора и три абортивные (справа)
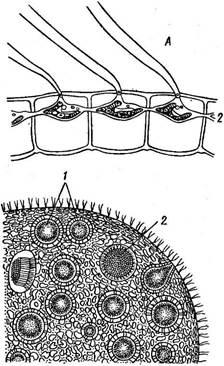
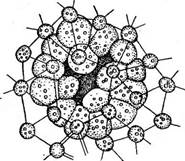 Рис. 76. УоЬох аигеив. Молодая дочерняя сфера с незамкнутым отверстием
Рис. 76. УоЬох аигеив. Молодая дочерняя сфера с незамкнутым отверстием
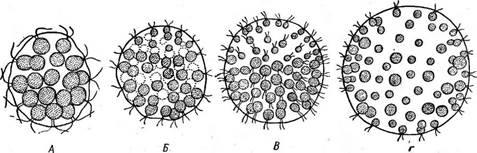 Рис. 78. Постепенная стерилизация клеток колоний у разных вольвоксовых. А — Еийо- ппа е1едапв; Б— Е. тШса; В — Р1еийоппа саИ[огтса; Г—Р. врНаегка. Более мелкие клетки чисто соматические, не способные к размножению
Рис. 78. Постепенная стерилизация клеток колоний у разных вольвоксовых. А — Еийо- ппа е1едапв; Б— Е. тШса; В — Р1еийоппа саИ[огтса; Г—Р. врНаегка. Более мелкие клетки чисто соматические, не способные к размножению