|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
КЛАСС КОНЪЮГАТЫ, ИЛИ СЦЕПЛЯНКИ, —
СОМШОАТОРНУСЕАЕ Относящиеся сюда одноклеточные и нитчатые зеленые водоросли отличаются полным отсутствием в жизненном цикле жгутиковых стадий, так как бесполого размножения с помощью зооспор у них нет, а половой процесс — конъюгация — заключается в слиянии амебоидных протопластов обычных вегетативных клеток. Помимо конъюгации широко распространено вегетативное размножение. Для подавляющего боль- шинства конъюгат характерен центральный, осевой хроматофор, париетальный хроматофор встречается сравнительно редко. Большинство пресноводные формы, некоторые живут на почве и в солоноватых водах. Класс включает четыре порядка, из которых главнейшие: мезоте- ниевые — Ме5о1аета1ез, зигнемо- вые — 2у^пеша1: а1е5 и десмидие- вые — БезгтйЫез. Для представителей первых двух порядков характерна цельная сплошная оболочка без пор, у десмидиевых клеточная оболочка состоит из двух или более частей и пронизана сложно устроенными порами. Порядок мезотениевые — Ме$о1аеп! а1е$ Одноклеточные формы. Клетки обычно цилиндрические, с закругленными концами, в поперечном сечении всегда округлые. Клетка одета сплошной гладкой оболочкой, поверх которой имеется слизистая обвертка. Одноядерные клетки содержат хроматофоры, различающиеся у разных родов, по форме. У рода мезотениум (Мево1аетит) хроматофор в виде осевой пластинки, с одним или несколькими пиреноидами; в зависимости от положения клетки хроматофор можно видеть или с его широкой стороны,, или в профиль (рис. 120). У родов цилиндроцистис (СуИпЛгосувИз) и нетриум (Ые1г1ит) в клетке имеется по два центральных в поперечном сечении звездчатых хроматофора, состоящих из массивной осевой части, содержащей пиреноид, и расходящихся по радиусам пластинок, у нетриума сильно изрезанных по краям (рис. 121, А).
У рода спиротения (8р[го1аегйа) хроматофоры париетальные, в виде спиральных лент с пиреноидами (рис. 122). Размножение мезотение- вых вегетативное, путем деления клеток в поперечной плоскости. Половой процесс — конъюгация. Конъюгирующие клетки, обычно погруженные в общую слизь, располагаются своими длинными осями или параллельно друг другу, или под прямым углом и в месте контакта образуют навстречу друг другу отростки. По мере удлинения отростков конъюгирующие индивиды несколько раздвигаются (рис. 121, Б). При растворении стенок в месте стыка отростков возникает конъюга- ционный канал, внутри которого встречаются и зливаются протопласты конъюгирующих клеток (рис. 121, В, Г). Зигота одевается толстой многослойной оболочкой и переходит в состояние покоя. При прорастании зиготы копуляционное диплоидное ядро делится дважды, причем число хромосом редуцируется. Как правило, все четыре гаплоидных ядра жизнеспособны. При прорастании зиготы, как правило, формируются четыре новых индивида. Однако у нетриума из четырех гаплоидных ядер обычно остаются жизнеспособными только два, и соответственно при прорастании зиготы развиваются только два проростка (реже четыре). Мезотениевые распространены в торфяных болотах, некоторые виды мезотениума и цилиндроцистиса встречаются на сырой земле, образуя слизистые скопления. Порядок зигнемовые — 2у^пеша(а1е$
Сюда относятся самые обычные нитчатые пресноводные водоросли. Все они, как правило, неветвящиеся, состоят из одного ряда цилиндрических клеток, одетых цельной оболочкой без пор и слизистым чехлом.
Благодаря чехлу нити зигнемовых и их скопления слизистые на ощупь в отличие от скоплений (тины), образованных другими нитчатками, например кладофорой. Центр клетки занят крупной вакуолью, цитоплазма занимает постенное положение. Хроматофоры у зигнемовых ' тех же трех типов, что и у мезотениевых. У рода мужоция (Мои^ео- {ш, рис. 123, Л, Б) — в виде плоской осевой пластинки с несколькими пиреноидами. Хроматофор мужоции своей широкой стороной обращен к свету, если же освещение чрезмерно, пластинка может повернуться на 90° и стать к свету в профиль. Такой поворот занимает около 30 мин. Два осевых звездчатых хроматофора, каждый с крупным центральным пиреноидом, наблюдаются у рода зигнема (1уцпета, рис. 124, А). Хроматофоры в виде одной или многих лент, расположенных. в постенной цитоплазме и опоясывающих клетку по спирали, характерны для наиболее широко распространенного рода спирогира {5р1го§уга, рис. 125, А). Лентовидные хроматофоры спирогиры обычно имеют неровные городчатые края, и у многих видов вдоль ленты посередине проходит гребень, вдающийся внутрь клетки, — темная полоса вдоль ленты. По средней линии хроматофора расположены пиреноиды, / окруженные крахмальными зернами. Одно ядро всегда расположено в центре клетки: у мужоции оно прижато к одной стороне хроматофора (рис. 123, Л), у зигнемы — между двумя хроматофорами (рис. 124, Л), а у спирогиры ядро, окруженное слоем цитоплазмы, подвешено на цито- плазматических тяжах, отходящих от постенной цитоплазмы, в центре вакуоли (рис. 125, Л). Любая клетка нити способна к росту и делению. После деления ядра образуется поперечная перегородка, разрезающая хроматофоры спирогиры пополам, у мужоции и зигнемы деление хро- матофоров предшествует формированию поперечной перегородки. Вегетативное размножение осуществляется благодаря разрыву нитей на отдельные фрагменты в результате отмирания промежуточных клеток, и даже отдельные клетки могут вырасти в новые нити. Половой процесс — конъюгация. Наиболее обычна так называемая лестничная конъюгация, которая происходит между клетками двух нитей. Нити располагаются параллельно друг другу и вначале склеиваются слизью. Затем противолежащие клетки образуют навстречу друг другу выросты, которые соприкасаются и срастаются своими концами. Удлиняясь, выросты постепенно раздвигают нити, так что возникает фигура в виде лестницы (рис. 123, В; 124, 5; 125, 5). Стенки на соприкасающихся концах отростков растворяются, и возникает узкий канал, соединяющий полости конъюгирующих клеток. " Оба протопласта, втянувшиеся в отростки, после растворения смежных стенок вступают в контакт. У всех видов спирогиры и у многих видов зигнемы протопласт одной из конъюгирующих клеток (отдающей) сокращается, отстает от стенок и постепенно проталкивается через канал в другую (воспринимающую) клетку, где и сливается с ее содержимым, образуя зиготу (рис. 124, 5; 125, 5, В). Сокращение протопласта обусловлено, по-види--- мому, тем, что жидкость из центральной вакуоли диффундирует в многочисленные пульсирующие вакуоли, появляющиеся в цитоплазме, которые выпрыскивают ее в пространство между протопластом и клеточной стенкой (рис. 125, 5). Появление сократительных вакуолей и отставание протопласта от стенки наблюдается и в воспринимающей клетке, но значительно позднее, чем в клетке, отдающей свое содержимое.
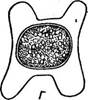
Помимо лестничной у тех же видов можно наблюдать иной тип конъюгации — боковой. При боковой конъюгации отростки, а затем Поскольку поведение конъюгирующих клеток у спирогир и многих зигнем несколько различно (воспринимающие клетки, более пассивные, можно назвать женскими, отдающие клетки, более активные, — мужскими), половой процесс в этих случаях можно определить как физиологически анизогамный, хотя морфологически он изогамен: конъюгирующие клетки морфологически одинаковы. У других видов зигнем и у всех видов мужоций слияние протопластов и образование зиготы происходит в конъюгационном канале (рис. 123, В), как у мезотение- вых. Здесь имеет место физиологическая изогамия, которая, как пра-~ вило, сопровождается морфологической изогамией. Описан лишь один вид мужоции, где конъюгирующие клетки и нити различались морфологически, т. е. наблюдалась морфологическая анизогамия, и у этого вида зигота всегда формировалась в конъюгационном канале. Образовавшаяся в результате слияния протопластов конъюгирующих клеток зигота округляется, выделяет толстую трехслойную оболочку и переходит в состояние покоя. В молодой зиготе еще можно различить хроматофоры обоих слившихся протопластов, позднее, в случае физиологической анизогамии, мужские хроматофоры разрушаются, остаются одни женские; при физиологической изогамии сохраняются все хроматофоры. Ядра сливаются незадолго до прорастания зиготы. При прорастании зигот зигнемовых (подробно изученном у спирогир А. Трендле, у многих видов зигнем — Л. И. Курсановым) происходит редукционное деление, причем из четырех гаплоидных ядер остается жизнеспособным только одно, соответственно развивается только один проросток (рис. 124, В; 125, Г). Порядок десмидиевые — Ое$гш(На1е$ Этот порядок, насчитывающий до нескольких тысяч видов, включает одноклеточные, реже нитчатые водоросли. Их клетки всегда состоят из двух симметричных половинок — полуклеток. У большинства родов в плоскости симметрии клетки имеется более или менее заметная перетяжка, исключение составляют род клостериум и некоторые виды рода пениум. Полуклетки могут быть самой разнообразной формы: цилиндрические с заостренными (клостериум), закругленными (пениум) или срезанными (плеуротениум) концами, а также уплощенные (космариум, эуаструм) или даже дисковидные (микрастериас). В разных положениях клетки выглядят неодинаково (см. рис. 127, 129, 131, 132). Полуклетки часто разделены более или менее глубокими вырезами на симметрично расположенные лопасти, углы их могут нести различные выросты. Клеточная стенка десмидиевых всегда состоит из двух половинок, имеющих скошенные края, плотно находящих друг на друга в плоскости симметрии клетки. В оптическом микроскопе стенка двухслойная, наружный слой часто импрегнирован соединениями железа, имеет желтоватую окраску и может нести шипы, бородавочки и другие выросты, расположенные определенным образом. Стенка пронизана сложно устроенными порами, часто группирующимися вокруг основания шипов, Рис. 126. С1с$1епит. А— вид клетки сбоку; Б — поперечный разрез клетки; В — выделение масс слизи с концов клетки; Г—конъюгация; Д, Е — конъюгация с образованием двойных зигот бородавок и других выростов стенки. Через поры выделяется слизь. Десмидиевые способны к слабому движению, находясь в контакте с субстратом. Движение обусловлено местным выделением слизи через специальные крупные поры, расположенные на концах клетки. В течение часа в этих местах могут выделиться неправильные червеобразные массы слизи, в два-три раза превышающие длину индивида (рис. 126, В). . Одно ядро находится в-цитоплазме в области перешейка, соединяющего две полуклетки. Хроматофоры в большинстве случаев осевые, по- одному в каждой клетке, хотя у многих видов космариума и некоторых других родов в каждой полуклетке может быть по два хроматофора. Такие осевые хлоропласты состоят из центральной — осевой части, у удлиненных форм, например у клостериума, имеющей палочковидную форму. От осевой части хлоропласта по радиусам расходится ряд пластинок или отростков. Пиреноиды обычно располагаются в центральной части хлоропласта. Реже хлоропласты постенные — париетальные, по- видимому, они произошли от центральных при расширении концов, радиально расходящихся отростков в париетальные лопастные пластинки и при редукции осевой части хлоропласта. У форм с массивными осевыми хлоропластами вакуоли небольших размеров. У клостериума и плеуротениума на концах клетки имеются терминальные вакуоли, содержащие кристаллы гипса, которые обнаруживают броуновское- движение. Размножение вегетативное — делением клетки в плоскости симметрии, половой процесс — конъюгация. Представителями одноклеточных десмидиевых могут служить роды клостериум (С1оз1епит), космариум (Созтапит), микрастериас (М1сгаз(епаз), стаураструм (51аигаз(гит) и эуаструм (Еиаз(гит). У клостериума (рис. 126, А) клетки веретеновидные, прямые или более или менее сильно изогнутые. Перетяжка в плоскости симметрии
отсутствует. Стенка состоит из двух половинок, спаянных в плоскости симметрии. В ней имеются поры, особенно крупные на концах клетки. Через поры выделяется слизь, приподнимающая конец клетки, этот конец перекидывается вниз, и слизь начинает выделяться с другого конца клетки, которая таким образом «кувыркается» (рис. 126, В)-, В плоскости симметрии клетки, куда обычно не заходит хлоропласт, находится одно крупное'ядро, часто различимое даже в живом состоянии. Каждая полуклетка содержит по одному крупному центральному хлоропласту, состоящему из осевого стержня, от которого по радиусам расходится несколько пластин. На поперечном разрезе хлоропласт имеет звездчатую форму (рис. 126, Б). На концах клетки находится по одной вакуоле с кристалликами гипса. У рода космариум (Созтапит, рис. 127, А, Б) клетки с глубокой перетяжкой, полуклетки разнообразной формы: округлые, пирамидальные, многоугольные. При рассматривании сверху клетка имеет эллиптические очертания, чем отличается от рода стаураструм (51аигаз1гит), клетка которого сверху выглядит треугольной или многоугольной (рис. 128). У рода микрастериас (ЛИсга81епаз) клетки плоские, с глубокой перетяжкой. Каждая полуклетка разделена на две боковые и одну полярную (срединную) лопасти. Боковые лопасти, как правило, глубоко надрезаны (рис. 129). У рода эуаструм (Еиа81гит) клетки большей частью удлиненной ■ формы, с глубокой перетяжкой, полуклетка с широкоокруглыми лопастями, имеет на конце выемку (рис; 130). Обычный способ вегетативного размножения десмидиевых — деле- Рис. 129. Мкга& епаз ние клетки на две в поперечной плоскости. Дочерние индивиды получают одну полуклетку от материнской клетки, а недостающую достраивают заново (рис. 127, В, Г).
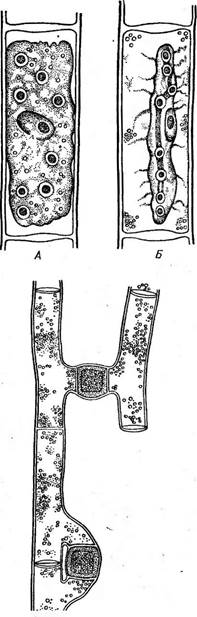
При конъюгации две клетки сближаются и одеваются общей слизью. У некоторых, например у клостериума, в области симметрии от каждой конъюгирующей клетки развиваются отростки, которые соединяются, образуя конъюгационный канал, внутри которого сливаются протопласты конъюгирующих клеток (рис. 126, Г). У многих десмидиевых половинки оболочек конъюгирующих клеток расходятся, протопласты освобождаются и сливаются (конъюгационный канал здесь не образуется). У некоторых видов (С1о51епит ИпеаШт и др.) протопласт каждой конъюгирующей клетки предварительно делится, формируя две гаметы, которые освобождаются и попарно сливаются. Таким образом возникают «двойные» зиготы (рис. 126, Д, Е). Зрелые зиготы обычно одеты трехслойной оболочкой. При прорастании зиготы копу- ляционное ядро редукционно делится и из четырех гаплоидных ядер, как правило, два остаются жизнеспособными. Таким образом, у десмидиевых из зиготы образуются два проростка, хотя есть и исключения (у гиалотеки только один проросток). Примерами нитчатых десмидиевых, у которых клетки при делении не разъединяются, а остаются плотно соединенными, могут служить роды десмидиум (ОезтШит) и гиалотека (НуаШНеса). Их клетки, как и у одноклеточных десмидиевых, состоят из двух симметричных полуклеток, в плоскости симметрии имеется неглубокая перетяжка. Нити обычно скручены по спирали и одеты более или менее мощным слизистым чехлом. У разных видов десмидиума клетки при рассмат- • I Заказ 552 ривании сверху имеют или овальные (Ю. суНпйпсит) или треугольные очертания (Л. зтаг(ги). Таким образом, у Л. $щ> аг{ги нить имеет форму трехгранной призмы, а у Л. суНпАпсигП — эллиптического цилиндра. Поскольку' нити скручены, выдающиеся ребра клеток у Л. зхюаг1ги выглядят в виде темно-зеленой полосы, идущей косо вдоль /^о^й нити (рис. 131). УЛ. суИпйпсит скручи- вание нити выражается в периодическом расширении и сужении ее в зависимости Ъхямы:. \ от того, широкой или узкой стороной по- ^г^гч вернута она к наблюдателю (рис. 132).
У гиалотеки (НуаШНеса) нить цилиндри-
Рис. 131. БеяпШит вшаг1гН. А — нить; Б— вид клетки сверху; В — конъюгация Рис. 132. БевтЫшт суИп- йгиит. А — нить; Б — вид клетки сверху чёская, клетки при рассматривании сверху имеют округлые очертания и отличаются очень слабой перетяжкой. Нить окружена мощным слизистым чехлом (рис. 133, А). У большинства нитчатых форм до конъюгации нить распадается на отдельные клетки (рис. 133, Б), однако у Б. 5Ы)аг1ги конъюгируют клетки, соединенные в нить. При конъюгации образуется конъюгационный канал (рис. 131, 6). * * * Отличительный признак конъюгат—полное отсутствие жгутиковых стадий и половой процесс в форме конъюгации. Прежде этим отличиям придавали большое значение и даже объединяли конъюгаты и диатомовые водоросли в особую, независимую от зеленых водорослей группу 2удорЬу1а. Однако отсутствйе жгутиковых стадий как вторичное явле- - ние наблюдается и среди других зеленых водорослей (например, у многих автоспоровых хлорококковых). В цитоплазме же конъюгирующих клеток ряда сцеплянок (нетриум, спирогира) появляются пульсирующие вакуоли, характерные для монадных жгутиковых клеток. Это позволяет рассматривать сливающиеся при конъюгации клетки как производные жгутиковых гамет других зеленых водорослей, причем появление пульсирующих вакуолей во время конъюгации можно считать атавизмом. Кроме того, слияние амебоидных гамет, характерное для конъюгат, было обнаружено у других зеленых водорослей, например у хламидомонады. В свете этих данных, а также учитывая сходство пигментов и продуктов запаса у конъюгат и остальных зеленых водорослей, невозможно сомневаться в принадлежности конъюгат к отделу зеленых водорослей. Их целесообразно выделить в особый класс как ветвь основного ствола рав- ножгутиковых, отделившуюся еще на уровне хламидомонад (у ряда видов которых имеется центральный хроматофор, столь типичный для подавляющего большинства конъюгат) и проделавшую далее длительную самостоятельную эволюцию.
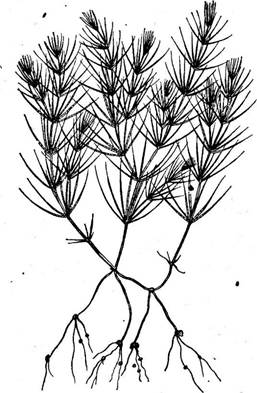
В пределах самого класса конъюгат простейшей группой являются мезотениевые с их просто организованной клеткой и, как правило, четырьмя жизнеспособными гаплоидными ядрами, возникающими в результате мейоза при прорастании зиготы. У них образуются четыре проростка. Из мезотениевых выводятся как десмидиевые, так и зигне^ мовые. Эволюция первых пошла по пути усложнения строения клетки, ' вторые, сохранив простые очертания клетки, эволюционировали в сторону образования нитчатых форм. И в том и в другом случае эволюция сопровождалась отмиранием части ядер, образующихся во время мейоза, и соответственно уменьшением количества проростков зиготы: у десмидиевых, как правило, до двух, у зигнемовых — до одного. КЛАСС ХАРОВЫЕ — СНАКОРНУСЕАЕ Класс харовые водоросли представлен всего одним порядком СЬага1е8 с тремя семействами, из которых два объединяют ископаемые формы и одно — СЬагасеае — наряду с ископаемыми включает и нынеживущие шесть родов. Наиболее обычны роды хара (СНага) и нигелла {N11е11а). От других зеленых водорослей они отличаются сложно устроенными многоклеточными половыми органами и расчленением нитчатого таллома. Нитчатые талломы харовых, всегда вертикально стоящие, достигают высоты около 20—30 см (иногда и метра). Мутовчатое ветвление придает им сходство с хвощами (рис. 134). Как неограниченно нарастающие оси («стебли»), так и боковые ветвй ограниченного роста («листья») дифференцированы на узлы и междоузлия. Узлы представляют собой клеточные пластинки. Они возникают из одной клетки в результате деления ее продольной перегородкой и затем отчленения продольными перегородками ряда (6—8) периферических клеток от двух центральных (рис. 135, Б). От периферических клеток узла «стебля» отделяются верхушечные клетки боковых ветвей ограниченного роста — «листьев», благодаря этому расположенных всегда мутовками. Узлы чередуются с междоузлиями, представленными у ни- теллы одной вытянутой (у самых крупных видов до 25 см) клеткой; у хары клетка междоузлия снаружи одета специальной корой.
Рост строго верхушечный: куполообразная апикальная клетка отчленяет перегородкой, параллельной ее плоскому основанию, сегменты. Каждый сегмент делится на двояковогнутую верхнюю клетку и двояковыпуклую нижнюю клетку (рис. 135, А). Двояковогнутая клетка дает начало узлу: она делится пополам продольной-перегородкой, после чего ряд изогнутых перегородок отделяет серию периферических клеток от двух центральных, позднее претерпевающих еще несколько делений (рис. 135, Б). Двояковыпуклые клетки не способны к делению, они только растут в длину, образуя междоузлия. Боковые ветви, или «листья», обнаруживают тот же способ роста, что и Рис. 134. СНага. Общий вид таллома главная ось, но клетки междоуз- И- Б В Рис. 135. СНага. А — продольный разрез конуса нарастания; Б — молодой узел в поперечном сечении; В — лопасти коры на молодом междоузлии стебля, состоящие из чередующихся узлов и междоузлий: 1 — верхушечная клетка, 2 — клетка-сегмент, 3, 7 — узел, 4, 6 — междоузлия, 5 — листья > лиев обычно остаются довольно короткими, а апикальная клетка сравнительно рано прекращает делиться, принимая вытянутую заостренную форму. От периферических клеток узлов, расположенных у основания «листьев», возникают ветви неограниченного роста — «стебель» ветвится. Базальные, или основные, узлы «листьев» Служат также местом возникновения характерной коры, свойственной большинству видов хары. Из некоторых клеток каждого базального узла возникают две нити: одна из них растет вверх, другая — вниз. Эти коровые нити плотно прилегают к клетке междоузлия и растут с той же скоростью, так что эта клетка с самого начала покрыта корой. Растущая вверх нить одного узла встречается с нитью, растущей вниз от вышерасположенного узла (рис. 135, В). Нити коры, одевающие стеблевые междоузлия, имеют то же строение и тот же способ роста, что и другие ветви растения. У них наблюдаются апикальный рост и расчленение на узлы и междоузлия (рис. 135, В). Узлы делятся двумя радиальными стенками на медианную и две латеральные клетки. Медианная клетка узлов коровых нитей делится тан- гентальной стенкой на более мелкую внутреннюю и более крупную наружную клетку, которая может дать или только маленький выступ, или развиться в шип; иногда она делится на ряд клеток. У многих видов хары «листья» также одеты корой, нити которой, однако, не дифференцированы на узлы и междоузлия (рис. 136, А). Прикрепляются талломы посредством многоклеточных разветвленных ризоидов, возникающих из периферических клеток нижнего узла главной оси. Рис. 136. СНага. А— половые органы на листе; Б, В, Г—разрез антеридия на разных стадиях развития; Д — манубриум, первичные и вторичные головки со сперматогенными нитями; Е — участок сперматогенной нити и сперматозоиды Клетки одеты целлюлозной оболочкой, в наружных слоях которой отлагается карбонат кальция. В постенной цитоплазме находятся многочисленные мелкие дисковидные хроматофоры, лишенные пиреноидов. В удлиненных клетках междоузлий хроматофоры расположены хорошо заметными продольными рядами в самом наружном слое цитоплазмы. Более глубокий слой цитоплазмы, граничащий с громадной вакуолью, обнаруживает интенсивное движение. В каждой клетке имеется восходящий ток цитоплазмы с одной стороны клетки и нисходящий — с другой. Линия раздела заметна по бокам клетки по бесцветной полосе, где нарушено плотное расположение хроматофоров и где стенка образует гребень, вдающийся внутрь клетки. Все клетки, способные к делению (т. е. апикальные, клетки-сегменты и клетки узлов), одноядерные, ядра делятся митотически. Не способные делиться клетки междо- уЗлиев, во взрослом состоянии многоядерные, содержат крупные ядра лопастной формы, размножающиеся амитотически. Бесполого размножения у харовых нет. Вегетативное размножение с помощью клубеньков, возникающих из нижних стеблевых узлов. Половой процесс оогамный. Половые органы имеют сложное строение. Как правило, оогонии и антеридии развиваются в непосредственной близости друг от друга, однако есть и двудомные виды. Оба вида половых органов формируются на вторичных боковых ветвях ограниченного роста, вырастающих из верхних узлов «листьев». При развитии антеридия боковая ветвь образует базальный узел, после чего апикальная клетка делится, формируя антеридий. Клетка базаль- ного узла антеридия на стороне, обращенной к оси, у однодомных форм- хары дает начало оогонию. Такйм образом, у хары оогоний направлен вверх, а антеридий — вниз (рис. 136, А), у однодомных видов нителлы оогоний образуется под антеридием. Апикальная клетка боковой ветви, из которой развивается антеридий, отчленяет одну или две дисковидные клетки у своего основания, принимает сферическую форму и делится двумя продольными и одной поперечной перегородками, образуя восемь клеток — октантов. Затем в каждом октанте последовательно возникают по две параллельные перегородки, так что получаются восемь диагональных рядов, каждый из трех клеток (рис. 136, Б, В). Восемь периферических клеток претерпевают плоскостной рост и развиваются в плоские изогнутые клетки — щитки, выпуклой стороной обращенные кнаружи. Их стенки имеют радиально расходящиеся складки, а содержимое в зрелом состоянии оранжево-красного цвета. Щитки составляют стенку шаровидного антеридия, которая у основания замыкается одной из дисковидных клеток, отчленившейся вначале и обычно вдающейся в полость антеридия (рис. 136, Г). Средние клетки значительно удлиняются в радиальном направлении, каждая образует клетку — рукоятку, которая таким образом отходит от центра щитка и несет на внутреннем конце шаровидную клетку — головку, развившуюся из самой внутренней клетки первоначального диагонального ряда. Каждая клет- ка-головка дает начало шести вторичным головкам, на которых развиваются длинные нити, обычно дважды-раздвоенные у основания. Эти сперматогенные нити заполняют полость антеридия плотным клубком. Каждая такая нить состоит из 100—200 дисковидных клеток, в которых образуется по одному спирально изогнутому двужгутиковому 1 сперматозоиду, сперматозоиды освобождаются при расхождении щитков антеридиальной стенки и ослизнении стенок клеток сперматогенных нитей (рис. 136, Д, Е). Клетка базального узла антеридия, которая развивается в оогоний, претерпевает два поперечных деления с образованием ряда из трех клеток. Самая верхняя из них дает оогоний, при этом она сильно увеличивается, самая нижняя — одноклеточную ножку, а средняя образует узел, который делится на центральную и пять периферических клеток. Последние вырастают в пять коровых нитей, которые уже на ранних стадиях окружают оогоний и вскоре отчленяют на своих концах одну (у хары) или две (у нителлы) клетки коронки. При дальнейшем развитии нити, не делясь, удлиняются и располагаются спирально. Клетки коронки мелкие, вертикально стоящие (рис. 136, А). В оогонии формируется одна яйцеклетка. Под коронкой, клетки которой плотно соединены, нити слегка разъединяются, так что образуется-щель, через которую проникает сперматозоид. Верхушка оогониальной стенки ослиз- няется, и сперматозоид оплодотворяет яйцеклетку. Вокруг оплодотворенной яйцеклетки выделяется целлюлозная оболочка, кнаружи находится стенка оогония и, кроме того, коровые нити. Внутренние стенкИ нитей утолщаются, опробковевают, в них может откладываться кремнезем^ а в полостях коровых нитей — обычно еще и известь. Таким образом, вокруг ооспоры образуется очень твердая обвертка и ооспоры переходят в состояние покоя. При прорастании ооспоры копуляционное ядро редукционно делится, формируются четыре гаплоидных ядра и появляется перегородка, делящая ооспору на две неравные части: 'верхняя клетка получает одно из четырех гаплоидных ядер, а три остальных ядра оказываются в нижней клетке, богатой запасными веществами. Эти три ядра затем дегенерируют. Стенка ооспоры разрывается на верхушке и верхняя клетка продольно, делится. Получившиеся две клетки растут в противоположных направлениях, образуя первый ризоид и вертикальную нить — предросток, или протонему, на которой развивается затем нормальный побег. По сложности строения вегетативных и половых органов, по наличию стадии протонемы, или предростка, при развитии зиготы даровые значительно отличаются от других зеленых водорослей, и ряд авторов рассматривают их не только как отдельный класс, но даже как самостоятельный отдел. Однако состав пигментов, наличие крахмала, гаплоидный характер (зиготическая редукция), относительно простое строение нителлы скорее свидетельствуют о том, что харовые водоросли представляют собой лишь очень специализированную ветвь зеленых водорослей. Харовые распространены преимущественно в пресных водах, хотя некоторые виды встречаются в солоноватых. Обычно они предпочитают водоемы, где нет сильных движений воды и где имеется песчаный или илистый грунт, в котором укореняются ризоиды. Харовые водоросли представляют немалый теоретический интерес: на них издавна изучали движение цитоплазмы. Кроме того, крупные клетки междоузлий, в которых цитоплазма ограничена постенным слоем, а центр клетки занят громадной вакуолью, служат объектами для исследования биоэлектрических явлений. Пользуясь в основном теми же методами и инструментами, которые применяли при изучении гигантского аксона кальмара, в клетке харовых водорослей удалось измерить разность потенциалов и сопротивление цитоплазматической мембраны. В состоянии покоя содержимое клетки заряжено отрицательно в основном за счет «хлористого насоса», который накачивает отрицательно заряженные ионы хлора из окружающей среды в центральную вакуоль. Мембрана в состоянии покоя относительно непроницаема для ионов кальция. Если раздражать клетку электрическим током, химическими веществами, уколом, то при раздражении, превышающем некоторый порог, мембрана в возбужденной области становится проницаемой для положительно заряженных ионов кальция: в результате их мгновенного поступления внутрь клетки мембрана разряжается. Электрический ток, возникший на этом участке, поднимает потенциал соседней области мембраны выше порогового, вызывая в ней такие же электрические изменения, которые распространяются, подобно волне, вдоль всей поверхности клетки. Таким образом, волну возбуждения, распространяющуюся по растительной клетке, можно вполне описать как потенциал действия и отнести ее к той же категории явлений, что и нервный импульс. Правда, имеются и различия: во-первых, растительная клетка реагирует медленнее, чем нервная, и, во-вторых, мембрана нервной клетки деполяризуется за счет мгновенного поступления внутрь клетки ионов натрия, а не кальция. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 480; Нарушение авторского права страницы

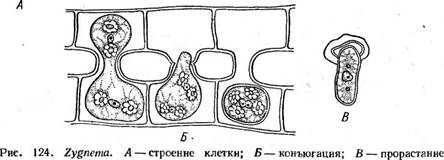 зиготы
зиготы
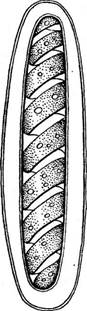
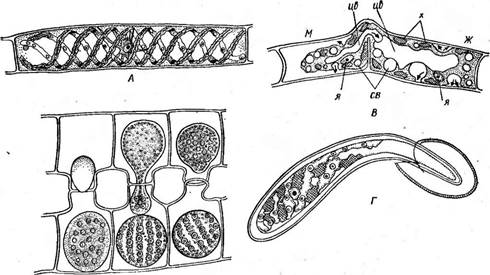 Рис. 125. 8рпо§уга. А — строение клетки; Б — конъюгация лестничная; В — конъюгация боковая'; Г — прорастание зиготы: Ж — женская гамета, М — мужская гамета; х— хлоропласт, я— ядро, цв — центральная вакуоль.
се — сократительные вакуоли
Рис. 125. 8рпо§уга. А — строение клетки; Б — конъюгация лестничная; В — конъюгация боковая'; Г — прорастание зиготы: Ж — женская гамета, М — мужская гамета; х— хлоропласт, я— ядро, цв — центральная вакуоль.
се — сократительные вакуоли

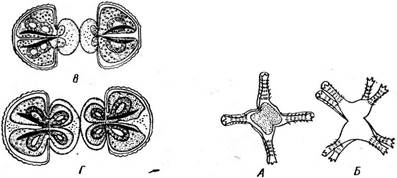
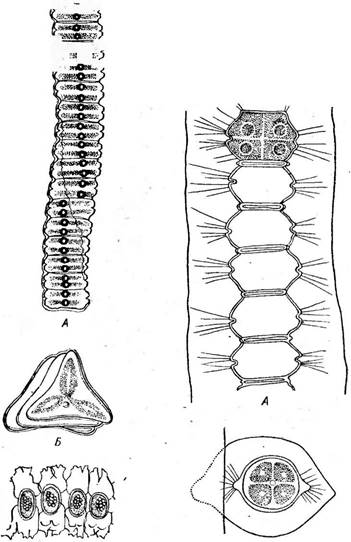
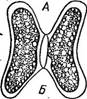
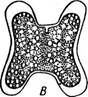
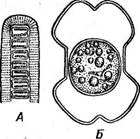 Рис. 133. НуаМНеса. А — нить; Б — конъюгат ция
Рис. 133. НуаМНеса. А — нить; Б — конъюгат ция