|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ОТДЕЛ КРАСНЫЕ ВОДОРОСЛИ — КНОООРНУТА
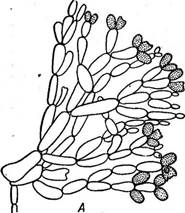
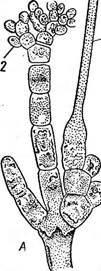
В хроматофорах красных водор'ослей помимо хлорофиллов («а» и «ё», но не «Ь») и каротиноидов (р-каротин, зеаксантин, антераксантин,. криптоксантин, лютеин, неоксантин) содержится еще ряд воднораство- римых пигментов — фикобилинов: фикоэритрины красного цвета, фико- цианины и аллофикоцианин синего цвета. От соотношения пигментов зависит окраска таллома, варьирующая от малиново-красной (преоб- 6 Заказ 552 Строение таллома красных водорослей довольно простое: известны даже одноклеточные коккоидные формы (пор- фиридиум), у многих талломы гетерот- рихальные и в виде разветвленных нитей, прикрепленных к субстрату с помощью ризоидов. Однако подавляющее большинство имеет псевдопаренхиматозные талломы, возникающие за счет переплетения боковых ветвей либо одной оси, неограни- Рис. 50. А — СЬап1гап81а с моно- ченно нарастающей с помощью верхушеч- спорами; Б — СаИННатгиоп с тет- ной клетки, либо многих таких осей; в распорангиями, нижний с тетра- первом случае говорят об одноосевом спорами (из них видны только строении, во втором — о многоосевом, или три), два верхних тетраспорангия у ' ' молодые (до образования спор) мультиосевом. Пластинчатые талломы истинно паренхиматозного строения, по- лучающиеся в результате как поперечных, .так и продольных делений клеток, встречаются среди красных водорослей редко (порфира).
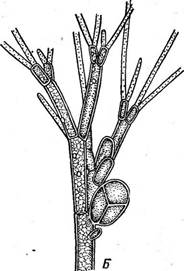
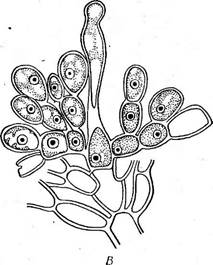
Клетка красных водорослей одета оболочкой, пектиновые и геми- целлюлозные компоненты которой сильно набухают и часто сливаются в общую слизь мягкой или хрящеватой консистенции, заключающую протопласты. Нередко в стенках откладывается известь. При делении клеток у большинства красных водорослей (класс флоридей) в стен- Рис. 51. А — ИетаНоп, ветвь с карпогоном (/)л антеридиями (2); Б—Ва1- гаскозрегтит, оплодотворенный карпогон; В — развитие карпоспор у Ва(га- сковрегтит ках остаются неутолщенные места — первичные поры, обычно отсутствующие у более примитивных красных водорослей (класс бангие- вые). Клетки одно- и многоядерные, хроматофоры, как правило, париетальные, многочисленные, в виде зерен или пластинок. У бангиевых встречаются звездчатые хроматофоры с центральными пиреноидами. Бесполое размножение осуществляется посредством неподвижных клеток, развивающихся из содержимого спорангия в числе одной — тогда их называют моноспорами или четырех — тогда говорят о тетраспорах (рис. 50). Тетраспоры формируются на диплоидных бесполых растениях — спорофитах (тетраспорофитах). В тетра- спорангиях перед образованием тетраспор происходит мейоз.
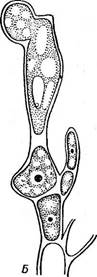
Половой процесс оогамный. Женский орган — карпогон (рис. 51, А) у большинства красных водорослей (флоридей) состоит из расширенной базальной части — брюшка, заключающего яйцеклетку, и отростка — трихогины (отсутствующей у бангиевых). Карпогон обычно развивается на особой короткой карпогон и альной ветви. Антеридии — мелкие бесцветные клетки, содержимое которых освобождается в виде мелких, голых, лишенных жгутиков мужских оплодотворяющих элементов — спермациев. Выпавшие из антеридиев спермации пассивно переносятся токами воды и прилипают к трихогине. В месте контакта спермация и трихогины их стенки растворяются и ядро спермация перемещается по трихогине в брюшную часть карпо- гона (рис. 51, Б), где сливается с женским ядром. После оплодотворе- 6* ' 83 Детали этого развития имеют важное систематическое значение. У одних красных водорослей содержимое зиготы (оплодотворенного карпогона) непосредственно делится с образованием неподвижных голых спор — карпоспор, у других из оплодотворенного карпогона (или из его дочерней клетки) вырастают ветвящиеся иити — гонимобласты, клетки которых превращаются в карпоспорангии, производящие по одной карпоспоре. Наконец, у большинства красных водорослей гонимобласты развиваются не непосредственно из брюшка оплодотворенного карпогона, а из особых вспомогательных — ауксилярных клеток. Последние могут быть удалены от карпогона или же располагаются на талломе в непосредственной близости от него. В тех случаях, когда ауксилярные клетки удалены от карпогона, из его брюшка после оплодотворения вырастают соединительные, или ообласте мные, нити. Их возникновению предшествуют митотические деления копуляционного ядра, и клетки ообластемных нитей содержат, таким образом, диплоидные ядра. Ообластемные нити подрастают к ауксилярным клеткам, в точке контакта оболочки растворяются и между клеткой ообластемной нити и ауксилярной клеткой устанавливается сообщение. Это слияние клеток не сопровождается слиянием их ядер (диплоидного ядра клетки ообластемной нити и гаплоидного ядра ауксилярной). Однако слияние с ауксилярной клеткой стимулирует деление диплоидного ядра клетки ообластемной нити и развитие гонимобластов, клетки которых содержат диплоидные ядра и продуцируют диплоидные карпоспоры. Гонимобласты, развивающие карпо- споры, обычно рассматриваются как особая генерация — карпоспо- р о ф и т. У наиболее высокоорганизованных красных водорослей ауксилярные клетки развиваются только после оплодотворения карпогона и в непосредственной от него близости. Совокупность ауксилярной клетки (или клеток) с карпогоном носит специальное название прокарпий. Здесь нет надобности в образовании длинных ообластемных нитей, связывающих карпогон и ауксилярные клетки, ауксилярная клетка просто сливается с брюшком оплодотворенного карпогона, после чего из нее развиваются гонимобласты с карпоспорами. Карпоспорангии часто располагаются тесными группами цистокарпиями, которые у многих представителей одеты псевдопаренхиматозной оболочкой, развивающейся из соседних с карпогоном клеток. Красные водоросли делят на два класса: бангиевые (Вап^юрЬу- сеае) и флоридеи (Р1опс1еорЬусеае). |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 306; Нарушение авторского права страницы