
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Порядок улотриксовые — 1Ло1ЬпсЬа1е8
Порядок объединяет зеленые водоросли, имеющие таллом в виде не- разветвленной нити, сложенной из одноядерных клеток. Реже таллом пластинчатый или трубчатый, но в онтогенезе развивается из нитчатого. Представителем может служить род улотрикс (ИШНпх) с наиболее известным видом V. гопа1а (рис. 94). Под Москвой эта водоросль встречается сравнительно редко в быстро текущих ручьях. Севернее, например в Неве, в прибойной полосе больших озер часто образует ватообразные обрастания на камнях. Нить состоит из одного ряда клеток, одетых' толстой оболочкой, под которой находится постенная цитоплазма, содержащая пластинчатый хроматофор, опоясывающий клетку изнутри в виде незамкнутого кольца. В хро- матофоре несколько пиреноидов, в цитоплазме одно ядро, центр клетки занят вакуолью. Все клетки нити одинаковые, за исключением базальной, которая вытягивается в короткий ризоид и служит для прикрепления нити к субстрату. Все клетки, за исключением базальной, способны делиться, обусловливая непрерывное нарастание таллома, а также образовывать репродуктивные клетки. При бесполом размножении во всех зеленых клетках развивается от двух до 16(32) четырехжгутиковых зооспор. Обычно этот процесс начинается в апикальной (конечной) клетке и продвигается к основанию нити.. Зооспоры, заключенные в слизистой обвертке, освобождаются через боковое отверстие в клеточной стенке. " Зооспоры несколько различаются по форме (от эллипсоидальных до шаровидных), содержат париетальный (пристенный) хлоропласт, не заходящий в переднюю часть зооспоры, имеют заметный глазок и один — семь пиреноидов. От переднего конца зооспоры отходят четыре жгутика. После периода движения (имеет место положительный фототаксис) зооспора останавливается, сбрасывая один за другим жгутики, прикрепляется боком к субстрату и прорастает в нить. При половом размножении в клетках возникают двужгутиковые гаметы в числе (4)—8—32(64). Гаметы выходят из клетки, заключенные в слизистый пузырь, в котором они двигаются, пока не освободятся. Половой процесс изогамный. У И.^гопа1а наблюдается гетероталлизм, т. е. копулируют гаметы из разных нитей, отличающихся в половом отношении («+» и «—»). В результате копуляции образуется сначала подвижная четырехжгутиковая зигота (планозигота), обнаруживающая отрицательный фототаксис. После некоторого периода движения оиа прикрепляется к субстрату и округляется, втягивая один за другим жгутики. При определенных условиях зигота прорастает в одноклеточный спорофит. Сначала она формирует трубковидный вырост, в который переходит ее содержимое. Этот вырост позднее вздувается, образуя на вершине грушевидные или дубинкообразные спорофиты, содержимое которых при созревании распадается на 4—16 (большей частью 8) четырехжгутиковых зооспор. Они выходят через отверстие, образующееся в результате ослиз- *нения части стенки спорофита, заключенные еще в слизистую обвертку, быстро исчезающую. Зооспоры оседают на субстрат и прорастают так же, как зооспоры, возникающие при бесполом размножении нитей уло- трикса. В менее благоприятных условиях улотрикс размножается вегетативно посредством фрагментации нитей. Схематически цикл развития улотрикса представлен на рис. 95. Другие виды улотрикса в неблагоприятных условиях переходят в пальмеллевидное состояние: клетки округляются, их стенки ослизня- ются, клетки расходятся и в таком состоянии делятся. При наступлении благоприятных условий каждая клетка такой пальмеллевидной стадии может превращаться в зооспору.
К порядку улотриксовые в качестве отдельного семейства относят ульвовые (Шуасеае), иногда их выделяют в особый порядок 1Л- уа1ез. Представители этой группы отличаются тем, что имеют не нитчатый, а пластинчатый таллом. Однако на ранних стадиях развития талломы их состоят из прикрепленной к субстрату нити, очень похожей на улотрикс, но рано или поздно в такой нити помимо поперечных делений- клеток наступают продольные, ведущие к формированию двухслойной пластинки. У рода ульва (Шуа, рис. 96) оба слоя клеток остаются плотно сомкнутыми, таллом достигает крупных размеров (кисти руки), имеет гофрированные края и прикрепляется к субстрату суженным в короткий черешок основанием. Клетки одноядерные, с по- стенным хроматофором. У близкого к ульве рода энтероморфа (Еп1е- готогрка) оба слоя клеток пластинки на ранней стадии разъединяются, и талломы, также крупные, принимают вид кишки или трубки с одно- слойной стенкой. Отсюда и русское название этой водоросли — кишеч- ница (рис. 97). Ульва и энтероморфа в основном морские водоросли, приуроченные к прибрежной (литоральной) зоне как в северных, так и в южных морях. Многие виды обоих родов выносят заметное опреснение воды и часто поднимаются в устья рек, а некоторые виды энтеро- морфы обычны и в пресноводных водоемах: часто встречаются в реках и прудах в Москве и под Москвой. Крупные талломы энтероморфы, развиваясь иногда в массовых количествах, а затем отмирая, портят воду, которая становится непригодной для пи-тья. Вегетативное размножение осуществляется отделившимися участками таллома. Бесполое размножение— зооспорами, как правило четырехжгутиковыми, которые образуются путем последовательного деления протопласта любой клетки на четыре — восемь частей. Половое размножение — двужгутиковыми гаметами. У разных видов обоих родов наблюдается изогамия или гетерогамия.
Оба рода детально изучены с точки зрения их цитологии и циклов развития. У этих родов зооспоры и гаметы никогда не формируются на ■ одном и том же экземпляре. Одни растения бесполые и несут только органы бесполого размножения — в их клетках развиваются только зооспоры. Другие растения половые, продуцируют только гаметы. Специальные исследования показали, что растения, образующие зооспоры, — спорофиты — имеют в ядрах диплоидный набор хромосом, являются диплоидными. При формировании Зооспор ядро редукционно делится и зооспоры, содержащие гаплоидные ядра, прорастая, дают гаплоидную генерацию — гаметофиты, которые продуцируют гаметы. Получившаяся в результате копуляции гамет зигота содержит одно диплоидное копуляционное ядро и прорастает сразу, без периода покоя, в диплоидную генерацию — спорофит. Таким образом, у ульвы и энтеро- морфы имеется правильное чередование генераций, или поколений: диплоидного спорофита и гаплоидного гаметофита. Оба поколения отличаются только цитологически (одно содержит 2п хромосом, - другое — п) и по органам размножения, которые на них развиваются. Внешне (морфологически) оба поколения друг от друга неотличимы. Такая смена поколений получила название изоморфной. И ульва, и энтероморфа — гетероталличные формы и их гаметофиты различны в половом отношении: одни продуцируют гаметы со знаком «+», другие — со знаком «—». Копуляция происходит, как и у улотрикса, лишь при встрече гамет разного знака. У гетерогамных видов талломы разного знака можно различить и невооруженным глазом по окраске плодущих участков: желтоватой у мужских и темно- зеленой у женских. Порядок хетофоровые — СЬае1ор1тога! е8 Сюда относят зеленые водоросли с гетеротрихальным строением, т-. е. имеющие нитчатые талломы, дифференцированные на распростертую по субстрату систему нитей, от которых отходят вертикально стоящие, более или менее разветвленные нити. Порядок включает ряд семейств. Семейство хетофоровые — СЬае{орЬогасеае. Центральный род семейства, у которого легко различимы обе части гетеротрихального таллома, — род стигеоклониум (5Н@еос1отит). Виды рода распространены в стоячей или медленно текущей воде на подводных предметах, на их верхней освещенной стороне. От нитей, стелющихся по субстрату, отходят вертикально стоящие ветвящиеся нити. Ветви обычно оканчиваются многоклеточными волосками из вытянутых бесцветных отмерших клеток. Строение клеток и размножение стигеоклониума в общем такое же, как у улотрикса (рис. 98). Исследования В. И. Успенской показали, что степень ветвления, образование волосков и другие особенности водоросли зависят от внешних условий. Так, ветвление стигеоклониума усиливается с повышением освещенности и с понижением концентрации нитратов в среде, развитие волосков также тормозится высоким содержанием в воде нитратов. При недостатке в воде азота и подщелачивании воды стигеоклониум легко переходит в пальмеллевидное состояние. При подщелачивании среды и отсутствий в ней свободной углекислоты стигеокло- -ниум формирует изогаметы (двух- или четырехжгутиковые — у разных видов). Наоборот, подкисление среды и присутствие свободной углекислоты способствуют образованию четырехжгутиковых зооспор. Род хетофора (СНае(оркога) похож на стигеок- лониум, но отличается от него тем, что его таллом погружен в плотную слизь. Полушаровидные или иной формы ярко-зеленые подуше.чки хетофоры часто встречаются в озерах на камнях и особенно на водных высших растениях. У разных видов стигеоклониума стелющаяся по субстрату система нитей может быть развита неодинаково: от рыхло разветвленной нити, часто состоящей из сравнительно немногих клеток, до компактного диска, образованного срастанием многочисленных ветвей. Развитие вертикальной системы обычно находится в обратной зависимости от развития распростертых нитей: у форм с хорошо развитой подошвой или диском вертикальная система может быть редуцирована до нескольких коротких веточек или представлена только волосками. Наоборот, при слабом развитии основания нижние клетки хорошо развитых вертикальных нитей дают ризоиды, способствующие прикреплению растения к субстрату. От форм, подобных стигеоклониуму, эволюция пошла в двух направлениях: в одних случаях эволюционировала восходящая часть таллома, а стелющаяся редуцировалась, в других случаях наблюдалась редукция вертикальной системы, а стелющаяся часть получала наибольшее развитие.
Первый путь эволюции привел к таким формам, как род драпарнальдия (БгарагпаШьа), у которого распростертая система практически отсутствует и таллом прикрепляется с помощью ризоидов, отходящих от нижних клеток восходящей системы. В отличие от большинства видов стигеоклониума у дра- парнальдии восходящая система четко дифференцирована на длинные и короткие ветви (рис. 99). Длинные, неограниченно нарастающие слабоветвя- щиеся оси образованы крупными, часто слегка бочонковидными бледными клетками со слаборазвитым хроматофором, который представлен экваториальным пояском с изрезанными краями. Короткие боковые ветви ограниченного роста, часто называемые ассимиляторами, растут пучками, на длинных осях. Они обильно ветвятся и состоят из коротких клеток с хорошо развитым постенным хроматофором, занимающим всю поверхность клетки. Концы этих ветвей часто превращаются в длинные многоклеточные волоски. Размножение драпарнальдии такое же, как и у стигеоклониума. Репродуктивные клетки (зооспоры, гаметы) образуются только в ассимиляторах, длинные оси -несут главным образом опорную функцию. 9 Заказ 552 У драпарнальдии, как и у стигеоклониума, общий габитус таллома может сильно изменяться в зависимости от условий среды. Свойственная драпарнальдии дифференцировка. таллома на длинные и короткие ветви подавляется высоким содержанием в среде связанного азота. Варьируя в среде концентрацию нитратов, В. И. Успенской (1930) удалось получить все переходы от типичной драпарнальдии к стн*геокло- ниумоподобным талломам. В СССР драпарнальдии встречается в быстро текущих ручьях и других местах с хорошей аэрацией. К. И. Мейер описал ряд видов драпарнальдии, выделенных позднее А. П. Скабичевским в отдельный род драпарнальдиелла (ОгарагпаШьеИа), которые эндемичны для оз. Байкал, где они образуют обширные заросли на глубине 10 м. Драпарнальдиелла имеет крупные, до 0, 5 м длины, талломы, как и у драпарнальдии, дифференцированные на длинные и короткие ветви. Однако в крупных клетках длинных осей хрома- тофор занимает всю поверхность в виде сеточки, а не в виде экваториального пояска, как у драпарнальдий. У ряда видов драпар- нальдиеллы из основания ассимиляторов развиваются коровые нити, плотно оплетающие осевую часть таллома и тем увеличивающие ее прочность. У драпарнальдиеллы помимо вегетативного имеется половое размножение. Копулируют гаметы, слегка различающиеся по размерам (гетерогамия), образованные в разных талломах (гетероталлизм). То, что у исследованных видов драпарнальдиеллы наблюдались только гаметы и никогда — зооспоры, заставляет предположить у них смену генераций: возможно, исследователи имели дело только с одной генерацией — гаметофитом. Это предположение тем более вероятно, что у некоторых видов стигеоклониума была установлена смена генераций (изоморфная и гетероморфная). Смена генераций наблюдается и у другого представителя этого семейства — у рода драпарнальдиоп- сиса фгарагпаШюрзьБ), у которого длинные оси состоят из чередующихся коротких и длинных клеток. Ассимиляторы отходят только от коротких клеток длинных осей. Как и у драпарнальдиеллы, от основания ассимиляторов отходят коровые нити, окутывающие длинные оси.
Изоморфная смена генераций установлена также у рода фричиел- ла (РгНзсЫеИа), единственный вид которого — /7. IиЬегоза найден до сих пор только в Африке, Индии и Японии, где он растет на сравнительно сухих почвах. Этот вид имеет сложный гетеротрихальный таллом, состоящий из подземной разветвленной системы нитей, которая дает первичные вертикальные нити. Эти нити прорастают сквозь тон-
кий слой почвы и развивают над ее поверхностью кустистые побеги (рис. 100). Другой эволюционный ряд форм характеризуется более или менее полной редукцией вертикальной системы. Примером таких большей частью эпифитных форм могут служить роды хетонема (Скае1опета) — редкий эпифит, обитающий в слизи разных пресноводных водорослей (тетраспоры, батрахоспермума), и афанохете (Аркапоскае1е) —обычный пресноводный эпифит на водорослях. У Аркапоскае(е вертикальная система представлена, как правило, только волосками (рис. 101); у Скае1опета — короткими нитями, несущими на конце одноклеточный волосок (рис. 102, А). Бесполое размножение—■ четырехжгутиковыми зооспорами, известен и половой процесс. У афанохете — резко выраженная гетерогамия. В крупных оогониях образуется по одной четырех- жгутиковой макрогамете (рис. 101, Б), которая выходит из оогония, непродолжительное время двигается, затем теряет жгутики, останавливается и оплодотворяется мелким четырехжгутиковым сперматозоидом (микрогаметой). Сперматозоид формируется в единственном числе в мелких, почти бесцветных антеридиальных клетках (рис. 101, 6). Половой процесс у хетонемы оогамный. Он был описан советским ботаником К. И. Мейером (1930) у Скае(опета 1ггеди1аге. У этого вида можно различить мужские и женские растения. Некоторые клетки мужских нитей превращаются в антеридии, их содержимое становится плотным и мелкозернистым, они несколько разрастаются, теряют хлорофилл и принимают желто-зеленую окраску, резко отличаясь от зеленых вегетативных клеток (рис. 102, А). В каждом антеридии образуется восемь сперматозоидов овальной формы с двумя жгутиками. Оогонии образуются на женских нитях, обычно на концах коротких боковых ветвей (рис. 102, Б). Все содержимое оогония превращается в одну яйцеклетку без жгутиков, которая выходит через отверстие в оболочке оогония (рис. 102, В). Оплодотворяется яйцеклетка вне оогония, т. е. имеет место примитивная оогамия, впервые описанная К. И. Мейером у зеленых водорослей — у Скае(опета ьгге^иШге. Позднее примитивная оогамия наблюдалась еще у нескольких зеленых водорослей из вольвоксовых и хлорококковых. Семейство трентеполиевые — Тгеп1ероНПасеае объединяет зеленые водоросли обычно гетеротрихальные, как правило, без волосков. Зооспорангии в отличие от большинства зеленых водорослей имеют иную форму по сравнению с вегетативными клетками. Половое размножение путем копуляции двужгутиковых изогамет. Род трентеполия (Тгеп(ерокИа, рис. 103, А) интересен тем. что приспособился к наземным условиям существования. Виды его особенно широко распространены в тропиках и субтропиках, где растут на камнях, скалах, стволах деревьев и как эпифиты на листьях. Некоторые входят в состав лишайников. В СССР талломы трентеполии образуют кирпично-красные или желтые налеты на стволах деревьев. У большинства видов таллом гетеротрихальный, дифференцированный на стелющиеся по субстрату нити и отходящие от них вертикальностоящие нити; у других видов вертикальная система нитей слабо развита. Более или менее шаровидные клетки толстостенные, протопласт без вакуоли, Четырехжгутиковые зооспоры образуются в специальных, отличающихся по форме от вегетативных клеток крючковидных спорангиях, сидящих на клетках-ножках (рис. 103, В). Эти спорангии легко отделяются и разносятся ветром в целом виде. Как показал К. И. Мейер, при созревании зооспорангии становятся многоядерньши, и стоит им попасть в каплю воды, как через несколько (3—5) минут содержимое их распадается на одноядерные участки, которые вырабатывают жгутики и превращаются в зооспоры. В шаровидных сидячих гаметангиях, которые также разносятся ветром в целом виде, образуются (в присутствии воды) двужгутиковые гаметы. Однако копуляция гамет наблюдается очень редко, гаметы большей частью развиваются партеногенетически, прорастая, как и зооспоры, на влажной коре в новые нити.
Семейство колеохетовые — Со1еосНае1асеае включает род колеохе- те (Со1еосНае1е)—пресноводный эпифит, обитающий главным образом на высших водных растениях. У одних видов типичный гетеротрихаль- ный таллом, погруженный в слизь, образует полусферические подушечки, у других видов произошла редукция восходящих нитей, и таллом представлен только распростертой по субстрату системой нитей. Последняя в свою очередь может состоять из рыхло расположенных разветвленных нитей или нити моуут срастаться боками, формируя компактный диск (рис. 104, А). Одноядерные клетки, включающие крупный постенный хроматофор с пиреноидом, обычно несут характерные, одетые воротничком щетинки, которые появляются на клетке в виде выступа, одетого сначала двухслойной стенкой. Затем наружный слой стен-
Рис. 104. Со1еосИае1е. А — внешний вид таллома С. ясиШа; Б — иити с антеридиями (слева) и оогониями (справа) у С- риШтЛа; В — прорастание ооспоры ки разрывается и остается в виде воротничка у основания щетинки, в которую развивается выступ, одетый только внутренней стенкой. Бесполое размножение колеохете осуществляется двужгутиковыми зооспорами. Половой процесс оогамный. Оогонии и антеридии развиваются обычно на одном талломе, хотя есть и двудомные виды (рис. 104, Б). В оогонии одна яйцеклетка, в антеридиях — маленьких бесцветных клетках — формируется по одному бесцветному двужгутиковому сперматозоиду. После оплодотворения ооспора одевается толстой оболочкой, а соседние с оогонием клетки дают ветвящиеся нити, которые, переплетаясь, образуют вокруг оогония и ооспоры псевдопаренхиматозную однослойную кору (рис. 104, В). Ооспора зимует и весной прорастает. При этом содержимое ее делится на 8—32 клетки, каждая из которых затем дает по одной зооспоре. Зооспора развивается в таллом. Первое деление копуляционного ядра в прорастающей ооспоре носит редукционный характер. Таким образом, растение колеохете гаплоидно, диплоидна только покоящаяся ооспора. Семейство плеврококковые — Р1еигососсасеае. Сюда относится род плеврококк (Р1еигососсиз) — повсеместно распространенная наземная водоросль, образующая зеленый порошистый налет на коре деревьев в нижней части стволов, на старых досках заборов, на стенах, камнях и т. п. Налет состоит из одиночных или собранных в группы по две, три или в пакетиках клеток в результате слабого разъединения продуктов деления (рис. 105). Плотная оболочка окружает протопласт без вакуолей, содержащий один париетальный хроматофор без пиреноидов. Единственный способ размножения — вегетативное деление клеток. В условиях повышенной влажности плеврококк образует короткие нити с немногими ветвями. На этом основании он рассматривается как редуцированный представитель порядка хетофоровых.
Из гетеротрихального таллома, свойственного хетофоровым, при подавлении развития той или иной его системы можно филогенетически вывести как распростертый гаметофит, так и вертикально стоящий спорофит высших растений. Среди хетофоровых, по-видимому, широко распространена смена, генераций, свойственная высшим растениям: она установлена у некоторых видов стигеоклониума, драпарнальдиопсиса, фричиел- лы; есть данные о приуроченности зооспорангиев и гаметангиев к разным индивидам трентеполии. Наконец, обилие среди представителей этого порядка наземных форм наряду со сходством пигментов и запасных продуктов побуждает именно среди хетофоровых искать предков высших растений.
Порядок эдогониевые — 0ес! одоп1а1е& Порядок включает нитчатые водоросли, одноядерные клетки которых делятся с образованием на их стенках особых «колпачков». По колпачкам эдогониевые водоросли легко отличить от всех остальных нитчатых водорослей. Кроме того, монадные клетки (зооспоры, сперматозоиды) эдогониевых в противоположность монадным клеткам других зеленых водорослей многожгутиковые: жгутики располагаются венном вокруг переднего бесцветного конца клетки. Такие монадные клетки называют стефаноконтными[11]. Базальные тела, или основания жгутиков, удерживаются фиброзным (волокнистым) кольцом, которое находится на границе бесцветной передней (апикальной) части — «головки» зооспоры (сперматозоида) и остальной части, занятой париетальным сетчатым хроматофором, — «тела». От фиброзного кольца между базальными телами, чередуясь с ними, отходят корни жгутиков: восходящие волокнистые корни в основном простираются в головку, а трубчатые нисходящие — в тело зооспоры (сперматозоида). Основания жгутиков и корни прочно соединены с фиброзным кольцом и сохраняются вместе с ним при обработках (необходимых для электронно-микроскопических наблюдений), при которых свободные части жгутиков утрачиваются. Это позволило более или менее точно подсчитать жгутики у монадных клеток эдого- ниевыУ водорослей, которые различаются только по числу жгутиков, что, несомненно, связано с размерами самих монадных клеток. Так, в зооспорах Оейо^отит (наиболее крупных) насчитывается около 120 жгутиков, в более мелких зооспорах Ви1Ъоскае(е жгутиков 37—49, а у сперматозоидов Оес1о^отит и Ви1ЬосНае1е — соответственно 30 и
Ряс. 106. Оейодопшт. Схема строения зооспоры по электронным микрофотографиям: Г —головка Зооспоры, Г —тело зооспоры, фк — фиброзное кольцо, ож — основания жгутиков ж — жгутик*, кзк- корни жгутиков, в —везикулы, наполненные слизистой субстанцией, д — дпктиосомы -Л—элементы эндоплазматической сети, и — митохондрии, х — хроматофор, ох — оболочи хро- матофора, г — тилакоиды, п — пиреноид, с - стигма, к - крахмал, я — ядро, р — рибосомы, им — ци- топлазматическая мембрана
Б, В, Г — деление клетки с образованием валика (1) и колпачка (2); Д — выход зоо- 1 споры; Е — нить с антеридиями (/) и сперматозоидами (2); Ж — оплодотворение яйцеклетки (4) в оогонии (3) сперматозоидом (2); 3—растение с оогониями (3) и мужскими карликовыми растеньицами — наннандриями (5) шесть — девять. Схема строения зооспоры Оейо^отит. представлена на рис. 105. Половой процесс оогамный. Представителем порядка может служить род эдогониум {Оейо^о- тит) — пресноводная водоросль, распространенная во всех частях света. Талломы ■ — в виде неветвящихся нитей, обычно в зрелом состоя- нии свободноплавающих, но в молодом состоянии прикрепленных к субстрату (к подводным высшим растениям, сваям и пр.) специальными базальными клетками. У видов, живущих в текучей воде, нити остаются прикрепленными всю жизнь. Цилиндрические клетки иногда вздуты в верхней части, одеты тонкой оболочкой, под которой располагается постенная цитоплазма, содержащая одно крупное, хорошо заметное даже в живом состоянии ядро и сетчатый цилиндрический хроматофор со многими пиреноидами (рис. 107, А). Центр клетки занят крупной вакуолью. Некоторые клетки нити делятся. До деления ядра в верхней части клетки под- поперечной перегородкой из внутренних слоев стенки возникает кольцевидное утолщение (складка) — валик, вдающийся в полость клетки. После деления ядра наружные слои стенки в области валика разрываются кольцевой трещиной, а сам валик начинает растягиваться (рис. 107, Б, В, Г) в длину, образуя новый цилиндрический участок стенки, вставляющийся между частями старой стенки. Часть протопласта с одним из дочерних ядер переходит в область растягивающегося валика и на уровне разрыва старой материнской стенки образуется поперечная перегородка. Таким образом стенка верхней из двух дочерних клеток в основном образована растянувшимся валиком, лишь сверху имеется несколько выступающая часть материнской стенки выше кольцевого разрыва первичной стенки. Она и образует то, что называют колечком или колпачком. Клетки с колпачком обычно делятся повторно и обнаруживают столько колпачков, сколько было делений (рис. 107, Б, В, Г). Бесполое размножение эдотониума осуществляется с помощью крупных зооспор, образующихся по одной из всего содержимого клетки (рис. 107, Д). Зооспора эллипсоидальная или шаровидная, темно- зеленая в задней части и с бесцветным носиком, окруженным целым венцом жгутиков на переднем конце. К. А. Гусева (1927) экспериментально доказала, что для формирования зооспор эдогониума имеет значение содержание в воде свободной углекислоты. Эти опыты позволяют объяснить неоднократно наблюдаемое разными авторами энергичное зооспорообразование эдогониума (да и других зеленых водорослей, например стигеоклониума, драпарнальдии) при перенесении из текучей воды в стоячую и в темноте. Именно в стоячей воде и в темноте возможно накопление углекислоты, выделяемой водорослью в процессе дыхания и стимулирующей зооспорообразование. Стенка клетки, в которой образовалась зооспора, разрывается около верхнего конца, обе ее половинки расходятся, и зооспора медленно выплывает, сначала одетая тонким слизистым пузырем, который вскоре исчезает (рис. 107, Д). После некоторого периода движения зооспора прикрепляется передним концом к субстрату, теряет жгутики, выделяет стенку и вытягивается в нить. Первое же деление молодого проростка обычно сопровождается формированием колпачка. Половое воспроизведение эдогониума оогамное. Оогонии — крупные, большей частью шаровидно или эллипсоидально вздутые клетки, содержащие по одной яйцеклетке (рис. 107, Ж). Антеридии — короткие дисковидные клетки с желтеющими редуцированными хроматофорами. В каждом антеридии развиваются по два желтоватых сперматозоида с венцом жгутиков на переднем конце (рис. 107, Е). Распределение половых органов у разных видов эдогониума неодинаково. Различают виды обоеполые, или однодомные, у которых мужские и женские органы развиваются на протяжении одной нити, и раздельнополые, или двудомные, когда оогонии и антеридии находятся на разных нитях. Большинство двудомных видов обнаруживает любопытный половой диморфизм. У этих — андроспоровых — видов оогонии формируются на обычных нитях, а. антеридии — на особых карликовых растеньицах. Последние берут начало из специальных андроспор, отличающихся от обычных зооспор мелкими размерами. Андроспоры развиваются или на тех же нитях, что и оогонии, или на отдельных нитях. Они оседают на оогонии или на соседние с ними клетки и прорастают, образуя двух- и трехкле- точное растеньице, верхние клетки которого функционируют как антеридии, давая по два сперматозоида (рис. 107, 3). В стенке зрелого оогония возникает отверстие, через которое сперматозоид проникает в оогоний и оплодотворяет яйцо (рис. 107, Ж, 3). После периода покоя ооспора прорастает с образованием четырех зооспор, которые дают начало новым растениям. При прорастании ооспоры имеет место мейоз, и таким образом вегетативная жизнь эдогониума проходит в гаплоидном состоянии. Другой род эдогониевых, также обычный в пресных водах, — буль- бохете (Ви1Ьос'Нае(е) — отличается тем, что нити обильно ветвятся и каждая клетка несет длинный бесцветный волосок со вздутым в виде луковицы основанием. Порядок бриопсидовые, или сифоновые, — Вгуор81< За1е$-§1рНопа1е5 Порядок объединяет водоросли, характеризующиеся сифоновым типом строения, отличительная черта' которого — отсутствие клеточных перегородок, возникающих, и то не всегда, только при основании органов размножения. Таллом часто крупных размеров и со значительной внешней дифференцировкой формально представляет собой одну клетку, одетую толстой, часто слоистой оболочкой, под которой находится постенный слой цитоплазмы, граничащий изнутри с непрерывной вакуолью. В цитоплазме находятся многочисленные дисковидные хроматофоры, которые наряду с хлорофиллами и каротиноидами, известными для других зеленых водорослей (а- и р-каротин, лютеин, виолаксантин, неоксантин), содержат два особых ксантофилла — сифонеин и сифоноксантин, встречающихся лишь у очень немногих других зеленых водорослей. Под хроматофорами в постенной цитоплазме, как правило, располагаются многочисленные ядра. Размножение вегетативное и половое. Бесполого размножения у большинства представителей нет. Половой процесс ге-, терогамный, реже изогамный. Большинство этих водорослей диплонты, редукционное деление ядра имеет место перед образованием гамет, у некоторых обнаружена смена генераций (изо- и гетероморфная). Подавляющее большинство бриопсидовых — обитатели тропических морей, лишь немногие растут в Черном море и севернее. Род бриопсис (Вгуор818, рис. 108) широко распространен в Чер Вегетативное -размножение бриопсиса легко осуществляется посредством отделения «перышек», которые в месте отделения закупориваются и могут развивать базальные ризоиды даже до отделения от материнского таллома, Кроме того, участки протопласта, вытекающие из таллома при его повреждениях, округляются и в течение нескольких минут формируют новые стенки. Затем они развиваются в новые растения бриопсиса. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 347; Нарушение авторского права страницы
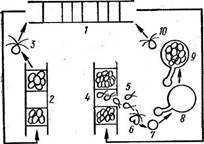 Рис. 95. Цикл развития Шо1кг1х гопа1а\
1 — вегетативная нить, 2 — нить с зооспо- рангиями, 3 — зооспоры. 4 — иить с гаме- тангиями, 5 —гаметы. 6 — слияние гамет (копуляция) с образованием зиготы (7), в — прорастание зигот с образованием спорофита, 9 — плодущий спорофит, 10 — зооспоры, образовавшиеся в спорофитах
Рис. 95. Цикл развития Шо1кг1х гопа1а\
1 — вегетативная нить, 2 — нить с зооспо- рангиями, 3 — зооспоры. 4 — иить с гаме- тангиями, 5 —гаметы. 6 — слияние гамет (копуляция) с образованием зиготы (7), в — прорастание зигот с образованием спорофита, 9 — плодущий спорофит, 10 — зооспоры, образовавшиеся в спорофитах
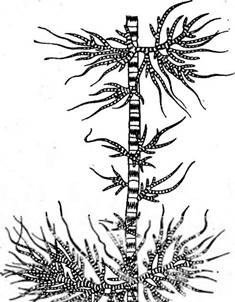

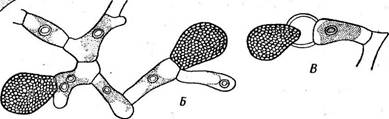 Рис. 102. С1гае1опета 1гге§и1аге. А —■ мужской талломе антернднямн; Б — женская нит-ь-с оогониями; В — выход яйцеклетки из оогония до оплодотворения
Рис. 102. С1гае1опета 1гге§и1аге. А —■ мужской талломе антернднямн; Б — женская нит-ь-с оогониями; В — выход яйцеклетки из оогония до оплодотворения

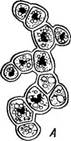

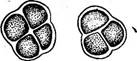
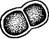
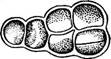 Рис. 105. Ркигососсиз. Общий вид
Рис. 105. Ркигососсиз. Общий вид
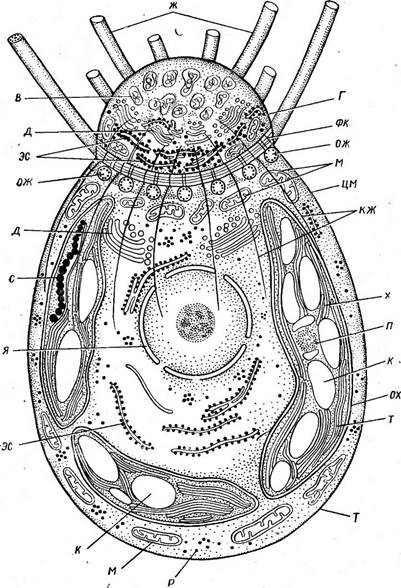
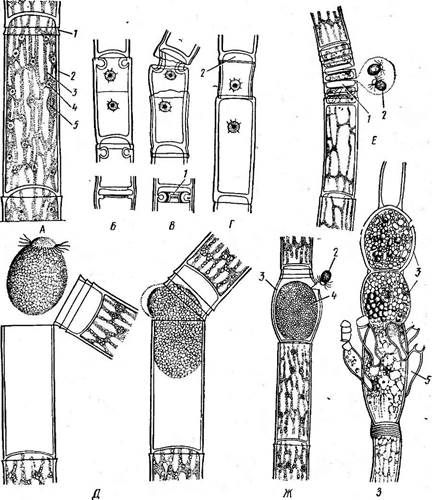 Рис. 107. ОейЩоп1ит. А— строение клетки:
/ — колпачок, 2 — стенка, 3— хлоропласт (хроматофор), 4 — пиреноид, 5 — ядро; Г
Рис. 107. ОейЩоп1ит. А— строение клетки:
/ — колпачок, 2 — стенка, 3— хлоропласт (хроматофор), 4 — пиреноид, 5 — ядро; Г