|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
ИЛИ ГОЛОСУМЧАТЫЕ,— НЕМ1А8СОМУСЕТШАЕ
Этот небольшой подкласс объединяет примитивные аскомицеты, у которых нет плодовых тел и сумки развиваются непосредственно на мицелии. Подкласс включает четыре порядка. Для порядка эндоми- цетовые (ЕпгёотусеЫев) характерно образование на мицелии одиночных сумок, развивающихся из зиготы без участия аскогенных гиф. У относящихся к этому порядку дрожжей мицелий отсутствует и сумки развиваются как одиночные свободные клетки. Смена ядерных фаз у эндомицетовых может быть разных типов. Сюда относятся как типичные гаплобионты, так и грибы с продолжительной диплоидной стадией (например, пекарские дрожжи — Засскаготусез се^еV^з^ае) и даже диплобионты (например, виды рода сахаромикодес — Засскаго- тусодез). Порядок тафриновые (ТарЬппа1е8) формально относится к подклассу гемиаскомицетов, так как его представители образуют сумки не в плодовых телах, а на мицелии, слоем под кутикулой растения- * хозяина. Однако тафриновые отличаются от остальных гемиаскомицетов тем, что в их цикле развития преобладает дикариотическая фаза, отсутствующая у других представителей этого подкласса. Кроме того, сумки у тафриновых не прототуникатные, как у остальных гемиаскомицетов, а эутуиикатные, как у высших аскомицетов. Поэтому многие микологи (Э. Гойман, Э. Мюллер и др.) считают эту группу вторично упрощенной, утратившей плодовые тела в результате приспособления к паразитизму. Кроме этих двух порядков к гемиаскомицетам относят порядки протомицетовые — Рго1ошусе1а1е8 и аскосферовые — А8созрЬаега1е8, положение которых в системе не ясно. Порядок эндомицетовые — Епйотусе1а1е$ У грибов из порядка эндомицетовые сумки расположены на мицелии поодиночке. У многих представителей этого порядка — дрожжей — настоящий мицелий отсутствует, клетки размножаются почкованием или, реже, делением, а сумки образуются как одиночные клетки. Большинство эндомицетовых — сапротрофы, обитающие на поверхности плодов и вегетативных частей растений, в нектаре цветков, в истечениях деревьев, в почвах. Паразитов в этой группе немного, например спермофтора хлопчатниковая — ЗрегторЫНога & оззури, развивающаяся в коробочках хлопчатника. Большое практическое значение имеют дрожжи — возбудители спиртового брожения и продуценты кормового белка. К эндомицетовым относятся и продуценты некоторых витаминов, например, широко используемый для производства рибофлавина эремотециум Эшби — Егето1кесшт азкЬу1. Эндомицетовые представляют большой теоретический интерес как возможное связующее звено между аскомицетами и их гипотетическими предками, близкими к современным зигомицетам. У представителей семейства диподасковые — Э1рос1а8сасеае образуется хорошо развитый мицелий, а на нем — цилиндрические многоспоровые сумки. Виды этого семейства обитают в истечениях растений, на древесине, в почве и не имеют практического значения. Однако это семейство наиболее интересно в теоретическом отношении, так как обнаруживает черты сходства с зигомицетами. В этом отношении наиболее детально изучен род диподаскус — йьройазсиз. У диподаскуса беловатого — й. а1Ыйиз, обитающего в истечениях различных растений, хорошо развит мицелий, состоящий из многоядерных клеток до 100 мкм длиной. На нем закладываются много- / ядерные гаметангйи, различающиеся по размеру. После слияния гаметангиев образовавшаяся зигота сразу же развивается в удлиненную многоспоровую сумку (рис. 236). Аскоспоры освобождаются из сумки пассивно. Они окружены слизистой обверткой, и " при ее набухании сумка растягивается и разрывается на вершине. Аскоспоры выходят из сумки и собираются на ее вершине в шарик, склеенный слизью. У другого вида—диподаскуса собранного— О. аццгед, п1и$ ' мицелий состоит из одноядерных клеток, и гаметангии также одноядерны. Этот гриб обитает в личиночных. ходах некоторых жуков-короедов. Развитие спор полового размножения у диподасковых имеет большое сходство с наблюдаемым у зигомицетов. Как и у зигомицетов, здесь сливаются многоядерные гаметангии (у О. а1Ыйи$), в процессе эволюции сменяющиеся одноядерными (у й. аццге^а1Ш) ■ Однако при слиянии многоядерных гаметангиев диподасковых множественная кариогамия отсутствует, а сливаются только два ядра. У некоторых зигомицетов (род эндогоне — Епдо^опе) при гаметангиогамии также сливаются только два ядра, а остальные ядра многоядерных гаметангиев дегенерируют. Сумку аскомицетов можно рассматривать как структуру, гомологичную зародышевому спорангию зигомицетов. У диподасковых их сходство увеличивается и тем, что количество спор в сумке может достигать нескольких десятков и строго не фиксировано, как у остальных аскомицетов.
Однако диподасковые существенно отличаются от зигомицетов тем, что зигота у них непосредственно развивается в сумку и не переходит в состояние покоя, а аскоспоры образуются по способу «свободного образования клеток», типичному для аскомицетов. Значительные различия существуют и в составе полисахаридов клеточных стенок у этих групп. У диподасковых, как и у других аскомицетов, в клеточных стенках имеются глюканы и хитин, но всегда отсутствует хитозан, характерный для зигомицетов.
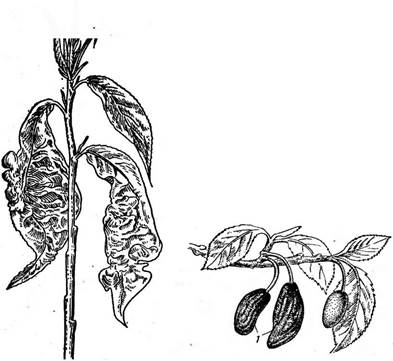
Для семейства эндомицетовые — Епйошусе1асеае в отличие от диподасковых характерно строго фиксированное число аскоспор в сумке (восемь или меньше восьми). К нему относятся преимущественно сапротрофы, обитающие на субстратах, богатых сахарами, однако отдельные виды паразитируют на грибах и животных. Некоторые эндомицетовые сбраживают сахара и используются в странах Азии и Африки для этой цели. Мицелий эндомицетовых состоит из хорошо выраженных гиф (род эремаскус — Егетазсиз), частично распадается на артроспоры (род эндомицес — Епйотусез) или почкуется (род эндомикопсис — Епйо- тусорзгз). У видов рода Егетазсиз, например у эремаскуса фертильного — Е. {ег1Ш5, гаметангиогамия сходна с таковой у диподасковых (см. рис. 234). Гаметангии представляют собой отростки клеток мицелия, содержащие по одному ядру. После их слияния зигота отделяется перегородками и превращается в восьмиспоровую сумку. У эндомицеса Магнуса — Епйотусез тадпизи, обитающего в соке деревьев, гаметангиогамия гетерогамная, т. е. гаметангии различаются по размеру и форме. У эндомикопсиса весеннего — Епйотусорзьз ьегпаИз, часто встречающегося весной в соке березы и других деревьев, сумки образуются апомиктически, из вегетативных клеток. Наиболее распространенная и практически важная группа из порядка эндомицетовые — дрожжи (семейство 5ассЬаготусе1асеае). Типичного мицелия у них нет, а есть лишь одиночные клетки, размножающиеся почкованием или делением (рис. 237, А). Если такие клетки не расходятся, формируется псевдомицелий (рис. 237, В). Половой процесс — копуляция двух вегетативных клеток (рис. 237, Г). Аскоспоры образуются в сумках, представляющих собой одиночные клетки. Предполагают, что дрожжи — организмы, вторично упрощенные в связи с обитанием в жидких сахаристых средах. Название «дрожжи» используется для группы грибов, которые на протяжении всего цикла развития или большей его части существуют в виде одиночных клеток, почкующихся или делящихся. Кроме грибов, образующих сумчатую стадию (семейство сахаромицетовые, объединяющее более половины известных родов дрожжей), к ней относят также грибы, филогенетически связанные с базидиомицетами. У таких дрожжей, например у видов из рода родоспоридиум — ЯоАозрогШшт, зигота прорастает дикариотическим мицелием, на котором развиваются телиоспоры. Они прорастают промицелием — короткими проростками, на которых экзогенно формируются гаплоидные споры, называемые обычно споридиями. Цикл развития у этих дрожжей имеет сходство с циклом развития головневых грибов (см. с. 445). Наконец, известны дрожжи, у которых половая стадия в цикле развития отсутствует (так называемые «аспорогенные дрожжи»). Их относят к классу дейтеромицеты, или несовершенные грибы, — Оеи1еготусе1е5. В отличие от двух предыдущих семейств, все представители которых типичные гаплобионты, сахаромицетовые объединяют грибы с разной продолжительностью гаплоидной и диплоидной фаз в цикле развития. У одних, например у видов из рода схизосахаромицес.— ЗсЫгозасскаготусез, наблюдается длительная гаплоидная фаза, а дип- лоидизация происходит непосредственно перед образованием сумки. Пекарские дрожжи — ЗассНаготусез сегеььз'ше после образования аскоспор в течение некоторого времени почкуются в гаплоидной фазе, после чего следует половой процесс — копуляция соматических клеток, и почкование продолжается уже в диплоидной фазе. При наступлении благоприятных условий (при хорошей аэрации и недостатке питания) такие диплоидные клетки превращаются в сумки. Наконец, среди са- харомицетовых есть грибы, у которых гаплоидная фаза сокращена до аскоспоры, а иногда наблюдается копуляция аскоспор уже в сумке. Примером таких диплобионтов может служить сахаромикодес Людвига — Засскаготусойез 1ис1~ощи. Дрожжи широко распространены на разнообразных субстратах, богатых сахарами: на поверхности плодов, в нектаре цветков, в истечениях деревьев и др. Представители некоторых родов, например липо- мицес — Ыротусез, обитают в почвах. Развиваясь на средах, содержащих сахара, дрожжи вызывают спиртовое брожение—превращение сахара в этиловый спирт и углекислый газ. Хотя этот процесс уже в древности использовался для приготовления алкогольных напитков и в хлебопечении, его причинная связь с развитием дрожжей была обнаружена лишь в 1876 г. Л. Пастером. Спиртовое брожение лежит в основе целого ряда пищевых производств— хлебопечения, виноделия, пивоварения, а также производства технического спирта из отходов целлюлозно-бумажной промышленности или мелассы. Однако известны многие виды дрожжей (например, виды из рода Ыротусез), не способные сбраживать сахара, а окисляющие их. Среди дрожжей, вызывающих спиртовое брожение, наибольшее значение имеют представители рода Засскаготусе5. Этот род объединяет как виды, встречающиеся в природе, так и «культурные» виды, представленные многочисленными производственными расами. Сахаромицеты способны к активному сбраживанию Сахаров и накоплению большого количества спирта (10—19 % по объему). Пекарские дрожжи — 5. сегеь'тае не выделяются из природных субстратов и существуют только в культуре. Они имеют слегка овальные клетки, размножающиеся почкованием. На поверхности клетки образуется сферический вырост, увеличивающийся в размерах и отделяющийся перетяжкой от материнской клетки. После отделения молодой клетки на ней и на материнской клетке остаются рубцы. По числу таких рубцов можно определить относительный возраст почкующейся клетки (иногда число их доходит до 30—40). В цикле развития пекарских дрожжей наблюдается довольно длительная диплоидная фаза. Сумки развиваются из диплоидных клеток и обычно содержат четыре аскоспоры (рис. 237, Б). Вид гетеротал- личен. Пекарские дрожжи представлены несколькими сотнями рас — винными, хлебопекарными, пивными и спиртовыми. Для получения спирта путем брожения используют картофель, зерно, патоку, а также сульфитные щелока — отходы деревообрабатывающей и целлюлозной промышленности. Это сырье предварительно осахаривают путем обработки солодом или кислотами, так как дрожжи не способны использовать полисахариды. Вина получают из виноградных и плодово-ягодных соков. В зависимости от степени использования сахара при брожении различают сухие, сладкие и крепленые вина. В процессе брожения кроме главных продуктов — этилового спирта и углекислого газа в небольших количествах образуются и другие вещества, от которых зависят специфический аромат и вкус вина. Разные вина получают путем использования определенных сортов винограда, рас дрожжей и соответствующей тех' нологии производства. Для производства пива используют охмеленное сусло из зерна яч^ меня, которое сбраживают дрожжами. Пиво содержит до 6 % спирта. Хлебопекарные расы дрожжей употребляют в хлебопечении и кондитерской промышленности. В тесте под действием дрожжей происходит спиртовое брожение и образующийся углекислый газ вызывает его подъем. В закваске ржаного теста кроме дрожжей содержатся молочнокислые бактерии, в нем идет смешанное брожение — молочнокислое, вызываемое бактериями, и спиртовое, вызываемое дрожжами. Поэтому в ржаном хлебе содержится молочная кислота, придающая ему характерный кисловатый вкус. Биомасса дрожжей, накапливающаяся в бродильных производствах в качестве отходов, содержит большое количество белков, углеводов и жиров и представляет ценный кормовой продукт. Некоторые виды дрожжей используют для производства кормовых белков. Кормовые дрожжи — СапсНйа иИИз хорошо растут на разнообразных средах, содержащих как гексозы, так и пентозы. Для их промышленного получения используют отходы различных производств — сульфитные щелока, гидролизаты древесины, мелассу и др. Биомасса кормовых дрожжей богата белками, содержащими все необходимые для животных аминокислоты. Некоторые виды рода Сап- (ПАа способны расти на углеводородах нефти, образуя биомассу, содержащую белки, жиры и другие ценные вещества. Дрожжи легко культивируются и быстро растут, поэтому их широко используют для изучения разнообразных процессов в биохимии, генетике и других областях биологии. На этих объектах проведены многочисленные исследования по локализации в клетке и регуляции активности ферментов, механизмам действия антибиотиков на грибную * клетку, цитоплазматической наследственности и многим другим вопросам. Эремотециум Эшби — Егето1кесшт азкЬуь— представитель небольшого семейства спермофторовые — ЗрегшорЬШогасеае — в природе встречается как паразит на коробочках хлопчатника. Его ярко-желтый мииелий состоит из многоядерных клеток. Окраска его обусловлена присутствием большого количества рибофлавина. На мицелии образуются продолговатые многоспоровые спорангии с веретеновидными спорами. Оболочка спорангия после созревания разрывается, споры освобождаются и прорастают. На появившихся коротких проростках развиваются сумки, содержащие от четырех до 32 веретеновидных аскоспор. Этот гриб используется в микробиологической промышленности для производства рибофлавина. Спермофторовые — группа низших аскомицетов, сочетающая примитивные признаки предков (например, образование спорангиев) с эволюционно продвинутыми признаками аскомицетов (клеточный мицелий, фиксированное число аскоспор в сумке). Порядок тафриновые — ТарЬппа1е$ / Порядок объединяет около 100 видов грибов — паразитов высших растений, вызывающих гипертрофию и деформацию пораженных органов. Сумки образуются непосредственно на мицелии плотным слоем, располагающимся под кутикулой пораженных органов растения. Плодовые тела отсутствуют. Дикариотический мицелий распространяется под эпидермисом, под кутикулой и по межклетникам в тканях растения. Мицелий однолетний или многолетний, зимующий в трещинах коры или почках растений. К порядку тафриновые относится один род тафрина — Таркппа, виды которого паразитируют на представителях разных семейств растений, но особенно часто на розоцветных, а также березовых, буковых, ивовых и др. Это высокоспециализированные паразиты, поражающие лишь определенные органы одного или близкородственных растений. Развиваясь на растениях, тафриновые вызывают у них разнообразные деформации пораженных органов: курчавость листьев (рис. 238), «дутые плоды», или «кармашки» (рис. 239), «ведьмины метлы» и галлы. Возникновение таких деформаций связано с тем, что тафриновые способны синтезировать фитогормоны, стимулирующие рост, — р-индолил- уксусную кислоту и вещества типа цитокининов, а также стимулиро Цикл развития тафриновых можно рассмотреть на примере тафри- ны деформирующей (Т. йе^огтапа)—возбудителя курчавости листьев « персика (рис. 238). Аскоспоры этого \ I гриба зимуют в трещинах коры и в \ I || д почках, где могут почковаться в гапло
идной фазе. Весной из них развивается дикариотический мицелий, заражающий листья. Дикарионтизация у этого вида происходит в результате деления ядра в почкующихся клетках. Пораженные листья увеличиваются в размерах, их листовые пластинки утолщаются, хотя жилки недоразвиваются и остаются укороченными. Такие листья имеют волнистую «курчавую» поверхность и желтую, розоватую, а позднее бурую окраску. Дикариотический ми- Рис. 238. Курчавость листьев персика, вызываемая ТарНппа Ле(огтап8
целий гриба распространяется по межклетникам в ткани листа, а на его гифах, расположенных под кутикулой на нижней поверхности листьев, развиваются сумки. В двухъядерных клетках гиф, расположенных между эпидермисом и кутикулой листа, происходит кариогамия. В верхней части аскогенной клетки образуется вырост, аскогенная клетка делится на две. Ядро в нижней клетке-ножке дегенерирует, а верхняя клетка развивается в сумку. В ней осуществляются мейоз и митотиче- ское деление ядер и вокруг восьми гаплоидных ядер формируются аскоспоры. Аскоспоры способны почковаться, поэтому сумки нередко бывают вторично многоспоровыми (рис. 240), Сумки образуют под кутикулой плотный слой. Созревая, они прорывают кутикулу листа и оказываются на его поверхности. Аскоспоры у тафрины активно выбрасываются из сумки. Рис. 240. Цикл развития Таркппа: / — почкование аскоспор. 2 — дикарионтизация. 3 — дикариотический мицелий в растении. 4 — слияние ядер дикариона, 5 — деление диплоидного ядра и образование материнской клетки сумки, 6, 7 — развитие сумки, 8 — слой сумок на поверхности пораженного органа растения У' некоторых тафриновых днкарионтизация происходит в результате копуляции аскоспор. У многих из них обнаружен гетероталлизм. Другой широко распространенный вид тафриновых — тафрина сливовая — Т. ргит поражает сливу, вишню, алычу, терн и черемуху, у которых она вызывает образование так называемых «дутых плодов», или «кармашков». У пораженных плодов разрастается перикарп (рис. 239), косточка не развивается, вместо ■ нее остается полость. У черемухи этот вид вызывает не только образование дутых плодов, но и махровость цветков, утолщение тычинок и чашелистиков. Мицелий у тафрины сливовой многолетний, он зимует в тканях побегов, а также в почках. Т. сегаы вызывает у вишни появление «ведьминых метел» — густых скоплений укороченных, сильно разветвленных побегов с мелкими, быстро опадающими листьями. Положение тафриновых в системе неясно. Хотя формально они должны относиться к гемиаскомицетам, так как не имеют плодовых тел, многие признаки сближают эту группу с эуаскомицетами. В их цикле развития наблюдается длительная дикариотическая фаза, а их сумки имеют эутуникатное строение и аскоспоры из них выбрасываются активно. Предполагают, что тафриновые филогенетически связаны с дискомицетами, а отсутствие у них плодовых тел — результат морфологического упрощения в процессе приспособления к паразитизму. |
Последнее изменение этой страницы: 2017-03-17; Просмотров: 267; Нарушение авторского права страницы

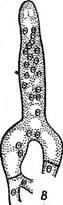
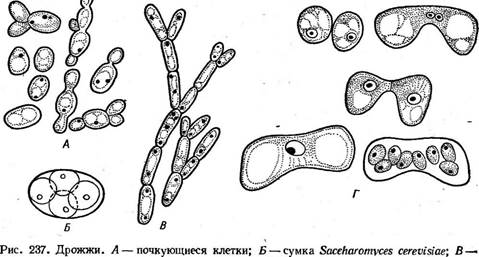 псевдомицелий; Г—половой процесс у Зскиовасскаготусев ос1оврогиз
псевдомицелий; Г—половой процесс у Зскиовасскаготусев ос1оврогиз