|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
II этап — скрещивание (гибридизация)
Бывает двух видов: 1. Близкородственное — инбридинг (позволяющее перевести рецессивные гены в гомозиготное состояние); 2. Неродственное (помогающее объединить в одном организме ценные признаки разных форм). III этап — отбор — завершающий этап селекции. Известно две формы отбора: · массовый (выделение группы особей, сходных по фенотипу, но дающих расщепление при размножении) · индивидуальный (выделение единичных ценных форм и раздельное выращивание Потомства каждой особи) приводит к созданию сорта или породы чистой линии. В селекции растений широко используется инбридинг, полиплоидия, искусственный мутагенез, отдаленная гибридизация. В области селекции растений много сделали известные селекционеры-генетики: И. В. Мичурин и Г. Д. Карнеченко, II. В. Цицин, П. II. Лукьяненко, В. Н. Ремесло, В. С. Пустовойт и л р. Ими были выведены высокоурожайные сорта сахарной свеклы, гречихи, хлопчатника; высокопродуктивные кубанские сорта пшеницы, украинские сорта «Мироновская-808, «Юбилейная-50», «Харьковская-63» и др. Селекция животных отличается от таковой у растении: животные дают мало потомков, у них позднее наступает половозрелость, они не размножаются вегетативно, отсутствует самооплодотворение. В селекции животных используют гибридизацию и отбор (массовый и индивидуальный), инбридинг и другие методы (М. Ф. Иванов, Н. С, Батурин и др.) Селекция микроорганизмов — молодая, развивающаяся отрасль селекции. Ее задача — получение высокопродуктивных микроорганизмов путем воздействия на исходные формы лучами Рентгена, ультрафиолетовыми лучами и химическими мутагенами. Чередование обработки мутагенами с отбором позволяет выделять штаммы, по продуктивности в десятки раз превосходящие исходный. Генетика Генетика популяций — наука о генетической структуре природных популяций и генетических процессах, происходящих в ней, таких, как дрейф генов, миграция, мутация и отбор. Все организмы состоят из больших популяций, в которых по законам генетики поддерживается равновесие генетического материала. Однако это равновесие постоянно нарушается мутационными процессами, миграциями, дрейфом генов и другими факторами. Все разнообразие в человеческих популяциях — есть результат мутационных изменений. Видный генетик С. С. Четвериков (1882—1959) внес существенный вклад в доказательство связи генетики с эволюцией. Он показал, что первые элементарные процессы начинаются в популяциях. Природные популяции при относительной фенотипичной однородности по генетической структуре разнородны и насыщены множеством открытых мутаций, образующих резерв {генетический груз) наследственной изменчивости. Под генетической структурой понимают соотношение в ней разных генотипов и аллельных генов. Английский математик Харди и немецкий врач Вайнберг установили, что при идеальных условиях — крупной популяции отсутствии мутаций, миграций и отбора — соотношение генотипов и аллельных генов во всех поколениях постоянно. Резерв наследственной изменчивости в популяции образуется за счет мутации. Доминантные мутации возникают редко, проявляются сразу и подвергаются отбору, Рецессивные мутации у гетерозиготных организмов фенотипически не проявляются, но при скрещивании насыщают генофонд популяции и образуют новые генотипы. Генофонд популяций пополняется также за счет генного потока — миграции особей из других популяций, приносящих новые гены. Они, также, как мутации, при скрещиваниях первое время у гетерозиготных организмов не проявляются. Одним из путей относительно быстрого изменения частот генов является случайное распределение генов, называемое дрейфом генов. Дрейф генов, случайная, нецеленаправленная смена частоты встречаемости аллелей в популяции, обуславливаемая периодическими популяционными волнами. Чаще всего дрейф генов встречается в малочисленных популяциях. В результате дрейфа генов в популяции может возрастать частота встречаемости редких аллелей, некоторые аллели могут исчезать; длительный период могут сохраняться мутантные аллели, что снижает приспособленность особей к условиям жизни. Резерв наследственной информации образуется еще и за счет комбинативной изменчивости, при которой в одном генотипе объединяются и обезвреживаются разнонаправленные мутации. Накапливаясь в популяции, скрытые мутации частично переходят в гомозиготное состояние и тогда проявляются фенотипически. В постоянных условиях стабилизирующий отбор (отбор в пользу нормы признака) устраняет их как несоответствующие условиям среды. В меняющихся условиях, при действии движущего отбора (отбора некоторых отклонений от установившейся нормы признаков), резерв наследственной изменчивости позволяет популяции приспосабливаться к новым условиям среды. Чем больше генотипов в популяции, тем шире ее норма реакции, тем вероятнее ее выживание в меняющихся условиях и возможность полнее использовать новые места обитания. Каждый биологический вид обладает уникальным генофондом, поэтому одной из важнейших задач человечества является охрана генофонда естественных популяций организмов.
102. Важным теоретическим обобщением исследований Н. И. Вавилова является разработанное им учение о гомологических рядах. Согласно сформулированному им закону гомологических рядов наследственной изменчивости, не только близкие в генетическом отношении виды, но и роды растений образуют гомологические ряды форм, т. е. в генетической изменчивости видов и родов существует определенный параллелизм. Близкие виды благодаря большому сходству их генотипов (почти одинаковому набору генов) обладают сходной наследственной изменчивостью. Если все известные вариации признаков у хорошо изученного вида расположить в определенном порядке, то и у других родственных видов можно обнаружить почти все те же вариации изменчивости признаков. Например, приблизительно одинакова изменчивость остистости колоса у мягкой, твердой пшеницы и ячменя. Трактовка Н.И.Вавилова. Виды и роды генетически близкие характеризуются сходными рядами наследственной изменчивости, с такой правильностью, что, зная ряд форм в пределах одного вида можно предвидеть нахождение параллельных форм у других видов и родов. Чем ближе родство, тем полнее сходство в рядах изменчивости. Значение закона 1. Закон гомологических рядов наследственной изменчивости позволяет находить нужные признаки и варианты в почти бесконечном многообразии форм различных видов как культурных растений и домашних животных, так и их диких родичей. 2. Он дает возможность успешно осуществлять поиск новых сортов культурных растений и пород домашних животных с теми или иными требуемыми признаками. В этом заключается огромное практическое значение закона для растениеводства, животноводства и селекции. 3. Его роль в географии культурных растений сопоставима с ролью Периодической системы элементов Д. И. Менделеева в химии. Применяя закон гомологических рядов, можно установить центр происхождения растений по родственным видам со сходными признаками и формами, которые развиваются, вероятно, в одной и той же географической и экологической обстановке.
103. Прямая репарация ДНК обеспечивает прямое восстановление исходной структуры ДНК или удаление повреждения. Широко распространенная система репарации такого рода — фотореактивация пиримидиновых димеров. Кроме нее, к этому типу относятся: репарация ДНК за счет 3'-5'-экзонуклеазной активности ДНК-полимеразы, репарация одноцепочечных разрывов ДН К с помощью полинуклеотиллигазы, а также генетическая репарация повреждений, вызванных алкильными или метильными группами, путем удаления этих групп специфическими ферментами.
Перед тем, как клетка делится, ее ДНК копируется (дублируется.) Две цепи молекулы ДНК имеют дополнительные пары оснований, последовательность нуклеотидов каждой пряди автоматически поставляет информацию, необходимую для создания своего партнера. Если две нити молекулы ДНК отделены, каждая из них может быть использована в качестве шаблона или шаблона для получения комплементарной цепи. Каждый шаблон и его новое дополнение образуют новую двойную спираль ДНК, идентичную оригиналу. Перед тем, как происходит репликация днк, ДНК должна быть раскручена. Кроме того, две нити должны быть разделены, так же, как две стороны молнии, разбивая слабые водородные связи, которые связывают парные основания. После того, как нити ДНК были размотаны, они должны быть ограждены, так что новые нуклеотидные партнеры могут водородных связей в них не найти. Фермент ДНК-полимераза затем перемещается вдоль открытой нити ДНК, присоединяя вновь прибывшие нуклеотиды к новой нити ДНК, комплементарной к шаблону.
Геномный импринтинг Геномный импринтинг — эпигенетический процесс, при котором экспрессия определённых генов осуществляется в зависимости от того, от какого родителя поступили аллели. Наследование признаков, определяемых импринтируемыми генами, происходит не по Менделю. Импринтинг осуществляется посредством метилирования ДНК в промоторах, в результате чего транскрипция гена блокируется. Обычно импринтируемые гены образуют кластеры в геноме. Импринтинг некоторых генов в составе генома показан для насекомых, млекопитающих и цветковых растений. Эпигенетика — в биологии, в частности в генетике — представляет собой изучение закономерностей эпигенетического наследования — изменения экспрессии генов или фенотипа клетки, вызванных механизмами, не затрагивающими последовательности ДНК.
№105 Механизмы эпигенетического регулирования экспрессии генов. 1. Эпигенетика – раздел в генетике, изучающий закономерности изменения экспрессии генов или фенотипа клетки, вызванных механизмами, не затрагивающими изменение первичной структуры ДНК и РНК. 2. Одним из основных эпигенетических механизмов является метилирование ДНК. А)Метилирование — это изменение молекулы ДНК путем присоединения метильной группы (-СH3) к нуклеотиду C, причем необходимо, чтобы за С следовал нуклеотид G. Последовательность нуклеотидов -CG- называется CpG динуклеотидом, или CpG сайтом. Метилирование происходит не во всех клетках одновременно, поэтому говорят о проценте метилирования определенного CpG сайта. Метилирование ДНК является одним из важных механизмов регулирования экспрессии генов. Показано, что с изменением профиля метилирования связаны такие заболевания, как различные виды рака, диабет первого и второго рода, шизофрения и т.д. Поэтому важно уметь анализировать профиль метилирования генома. Б)Ремоделирование хроматина Эпигенетические факторы влияют на активность экспрессии определенных генов на нескольких уровнях, что приводит к изменению фенотипа клетки или организма. Одним из механизмов такого влияния является ремоделирование хроматина. Хроматин — это комплекс ДНК с белками, прежде всего, с белками-гистонами. Гистоны формируют нуклеосому, вокруг которой накручивается ДНК, в результате чего обеспечивается её компактизация в ядре. От густоты расположения нуклеосом в активно экспрессирующихся участках генома зависит интенсивность экспрессии генов. Хроматин, свободный от нуклеосом, называется открытым хроматином. Ремоделирование хроматина — это процесс активного изменения «густоты» нуклеосом и сродства гистонов с ДНК. В) Модификации гистонов Хотя модификации аминокислот в гистонах происходят на всей молекуле белка, модификации N-хвостов происходит значительно чаще. Эти модификации включают: фосфорилирование, убиквитилирование, ацетилирование, метилирование, сумоилирование. Ацетилирование является наиболее изученной модификацией гистонов. Так, ацетилирование ацетилтрансферазой 14-го и 9-го лизинов гистона H3 (H3K14ac и H3K9ac, соответственно) коррелирует с транскрипционной активностью в данном районе хромосомы. Это происходит из-за того, что ацетилирование лизина меняет его положительный заряд на нейтральный, что делает невозможным его связь с негативно заряженными фосфатными группами в ДНК. В результате, происходит отсоединение гистонов от ДНК, что приводит к посадке на «голую» ДНК комплекса SWI/SNF и других транскрипционных факторов которые запускают транскрипцию. Это «цис» - модель эпигенетического регулирования. Гистоны способны поддерживать своё модифицированное состояние и выступать матрицей для модификации новых гистонов, которые связываются с ДНК после репликации.
106. Уровни организации хроматина. Краткий ответ: Первый уровень укладки ДНК в хроматине обеспечивается нуклеосомами. Они представляют собой округлые частицы диаметром 15 нм, которые связаны между собой участками ДНК длиной около 20 нм. Отдельная нуклеосома состоит их белковой сердцевины, на которую накручена молекула ДНК. Второй уровень укладки ДНК обеспечивается взаимодействием линкерной ДНК с гистоном H1. Молекула гистона H1 своим глобулярным доменом связывается с двумя витками ДНК на нуклеосоме. Одновременно C-концевой фибриллярный домен гистона H1 вступает в контакт с линкерной ДНК. В результате соседние нуклеосомы приближаются друг к другу, формируя группы из 6-8 частиц – нуклеомеры ( супербусины) диаметром 25-30 нм. Третий уровень укладки ДНК представлен хроматиновыми фибриллами диаметром 30 нм, которые хорошо видны в электронном микроскопе в интерфазных ядрах и митотических хромосомах. Они имеют суперспиральную структуру и содержат максимально сближенные между собой нуклеомеры. Четвертый уровень укладки ДНК обеспечивается взаимодействием фибрилл диаметром 30 нм с ядерным матриксом. При этом формируются петлевые домены, содержащие в среднем 90 тысяч пар нуклеотидов. В расправленном состоянии их длина может достигать 20 мкм. Пятый уровень укладки ДНК связан с формированием групп из 18-20 петлевых доменов, прикрепленных в виде розетки к общему центру из белков ядерного матрикса. Розетки из петлевых доменов находятся в хроматине в компактном состоянии, образуя округлые гранулы диаметром около 150 нм – хромомеры. Шестой уровень укладки ДНК определяется формированием хромонемы – фибриллярной структуры диаметром 200-300 нм, состоящей из плотно упакованных хромомеров.В хроматине интерфазного ядра хромонемы обычно не выявляются. Они становятся видимыми только при конденсации хроматина в профазе митоза, а также в ранней телофазе при деконденсации хромосом. Седьмой уровень укладки ДНК состоит в образовании хроматид (однохроматидных хромосом ) из хромонем. Толщина хроматиды составляет в среднем 700-800 нм. Если учесть, что толщина хромонемы обычно равна 100-200 нм, то коэффициент упаковки для хромосомного уровня составляет не более 10. Способ укладки хромонемы в хромосоме изучен недостаточно. У одних видов хромонема имеет вид спирали, у других в одной хромосоме могут обнаруживаться две и более параллельные друг другу хромонемы. Хромосомный уровень укладки ДНК в большей степени, чем другие уровни, отражает видовые особенности организации генома эукариот. Полный ответ: Молекулы ДНК имеют диаметр 2 нм, но их длина в хромосомах может достигать нескольких сантиметров. Очевидно, что упаковка таких длинных молекул в объеме клеточного ядра, имеющего диаметр всего 5-8 мкм, должна быть в высшей степени регулярной. Проблема укладки молекул ДНК в ограниченном объеме ядра осложняется еще и тем, что одновременно необходимо обеспечить возможность локальной распаковки ДНК и доступа к ней ферментов репликации и транскрипции. Вот почему хроматин в клеточном ядре образует сложные пространственные структуры с несколькими уровнями организации. Первый уровень укладки ДНК в хроматине обеспечивается нуклеосомами. Они представляют собой округлые частицы диаметром 15 нм, которые связаны между собой участками ДНК длиной около 20 нм. Отдельная нуклеосома состоит их белковой сердцевины, на которую накручена молекула ДНК. Белковая сердцевина нуклеосомы, или кор, имеет форму диска диаметром 11 нм и толщиной 6 нм. Она содержит по две молекулы гистонов H2A, H2B, H3 и H4. Если развернуть сердцевину, то можно обнаружить, что молекулы гистонов соединены в последовательности H2A, H2B, H4, H3, H3, H4, H2B, H2A. При сворачивании сердцевины молекулы гистонов располагаются как бы в два этажа, наподобие винтовой лестницы. Молекула ДНК в виде левозакрученной суперспирали совершает 1, 75 оборота вокруг сердцевины. При этом в непосредственный контакт с гистонами вступает 146 пар нуклеотидов. Длина линкерной ДНК, соединяющей соседние нуклеосомы, колеблется в пределах 40-70 пар нуклеотидов в зависимости от типа клетки. Таким образом, на одну нуклеосому приходится в среднем около 200 пар нуклеотидов ДНК. Нуклеосомы укорачивают молекулу ДНК примерно в 7 раз. Они обнаружены во всех эукариотических клетках и даже у ДНК-содержащих вирусов. Однако это не означает, что вся ДНК клеточного ядра связана с нуклеосомами, и некодирующие повторы могут иметь другую укладку. Второй уровень укладки ДНК обеспечивается взаимодействием линкерной ДНК с гистоном H1. Молекула гистона H1 своим глобулярным доменом связывается с двумя витками ДНК на нуклеосоме. Одновременно C-концевой фибриллярный домен гистона H1 вступает в контакт с линкерной ДНК. В результате соседние нуклеосомы приближаются друг к другу, формируя группы из 6-8 частиц – нуклеомеры ( супербусины) диаметром 25-30 нм. Последовательность аминокислот в C-домене гистона H1 гомологична первичной структуре некоторых регуляторов транскрипции. В связи с этим предполагается, что гистон H1 конкурирует с факторами транскрипции за связывание с линкерной ДНК, контролируя тем самым активность генов. Третий уровень укладки ДНК представлен хроматиновыми фибриллами диаметром 30 нм, которые хорошо видны в электронном микроскопе в интерфазных ядрах и митотических хромосомах. Они имеют суперспиральную структуру и содержат максимально сближенные между собой нуклеомеры. В формировании фибрилл диаметром 30 нм принимают участие гистон H1, а также негистоновые белки с HMG-доменом. За конденсацию хроматина в фибриллу диаметром 30 нм отвечает, прежде всего, C-концевой домен гистона H1. При этом первостепенное значение приобретает уже не взаимодействие гистона с ДНК, а взаимные связи гистоновых молекул между собой. Образование нуклеомеров и хроматиновых нитей диаметром 30 нм вызывает дальнейшую компактизацию ДНК в 40-50 раз. Четвертый уровень укладки ДНК обеспечивается взаимодействием фибрилл диаметром 30 нм с ядерным матриксом. При этом формируются петлевые домены, содержащие в среднем 90 тысяч пар нуклеотидов. В расправленном состоянии их длина может достигать 20 мкм. Концы таких петель прикреплены к ядерному матриксу в особых точках, обозначаемых как MARs (matrix attachment regions) или SARs (scaffold attachment regions). Эти точки содержат молекулы свободной ДНК (“форум-ДНК”) длиной 50-150 пар нуклеотидов, которые устойчивы к действию нуклеаз и выделяются из ядра независимо от высокомолекулярной ДНК. Точки MARs содержат также топоизомеразу II, участвующую в формировании изгибов ДНК. Со стороны ядерного матрикса прикрепление хроматиновых нитей обеспечивается ламином A. Петлевые домены являются типовой структурно-функциональной единицей хроматина. Домен во многом автономен, независимо реплицируется и транскрибируется. В своем составе домены имеют кластеры генов, которые связаны функционально. Петлевые домены обеспечивают компактизацию молекулы ДНК в 700 раз. Пятый уровень укладки ДНК связан с формированием групп из 18-20 петлевых доменов, прикрепленных в виде розетки к общему центру из белков ядерного матрикса. Розетки из петлевых доменов находятся в хроматине в компактном состоянии, образуя округлые гранулы диаметром около 150 нм – хромомеры. При активации локализованных в хромомере генов его величина может возрастать до 300 нм и более. Шестой уровень укладки ДНК определяется формированием хромонемы – фибриллярной структуры диаметром 200-300 нм, состоящей из плотно упакованных хромомеров.В хроматине интерфазного ядра хромонемы обычно не выявляются. Они становятся видимыми только при конденсации хроматина в профазе митоза, а также в ранней телофазе при деконденсации хромосом. Хромомерный и хромонемный уровни укладки позволяют укоротить длину молекулы ДНК в 10 000 раз. Седьмой уровень укладки ДНК состоит в образовании хроматид (однохроматидных хромосом ) из хромонем. Толщина хроматиды составляет в среднем 700-800 нм. Если учесть, что толщина хромонемы обычно равна 100-200 нм, то коэффициент упаковки для хромосомного уровня составляет не более 10. Способ укладки хромонемы в хромосоме изучен недостаточно. У одних видов хромонема имеет вид спирали, у других в одной хромосоме могут обнаруживаться две и более параллельные друг другу хромонемы. Хромосомный уровень укладки ДНК в большей степени, чем другие уровни, отражает видовые особенности организации генома эукариот. Каждый вид, как известно, имеет характерный для него набор хромосом. Однако хромосомы можно наблюдать в микроскоп только при делении клеток митозом или мейозом. В ядрах неделящихся клеток хромосомы находятся в деконденсированном состоянии, образуя хроматин. Тем не менее, даже в интерфазном ядре хромосомы сохраняют свою индивидуальность, занимая в нем определенные хромосомные территории. Согласно К. Раблю (1885) хромосомы прикреплены к нуклеолемме теломерными концами, тогда как центромерные участки располагаются ближе к центру ядра. Новейшие исследования интерфазного ядра с помощью конфокальной микроскопии подтверждают эту концепцию, добавляя к ней ряд существенных деталей. В частности, гомологичные хромосомы локализованы на противоположных сторонах ядра. При активации генов и сопутствующей этому деконденсации хромосомы удлиняются, смещаясь к центру ядра. Вот почему в животных и некоторых растительных клетках гетерохроматин концентрируется преимущественно по периферии, тогда как эухроматин занимает центральную область ядра.
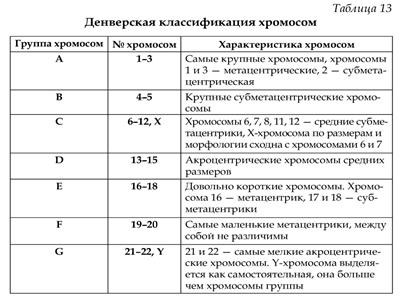
ВОПРОС № 107 Принцип, лежащий в основе Международной Денверской классификации хромосом человека Принципы: 1) различия в длине хромосом 2) расположение центромеры Хромосомы располагаются в порядке уменьшения их длины. Все пары хромосом распределены на 7 групп и пронумерованы арабскими цифрами: группа А(1-3) – большие метацентрические, В(4-5) – большие субметацентрические, С(6-12) – средние субметацентрические и Х-хромосома, D(13-15) – средние акроцентрические, Е(16-18) – небольшие мета- и субцентрические, F(19- 20) – самые короткие метацентрические, G(21-22) – самые мелкие акроцентрические, Y-хромосома. Центромерный индекс отражает длину короткого плеча к длине всей хромосомы. ОСНОВНАЯ ЧАСТЬ Основы существующей унифицированной классификации хромосом были заложены в 1960 году в Денвере. В основу классификации положены различия в длине хромосом и расположении центромеры. На основании различий в длине выделены 23 пары хромосом, при этом парам, имеющим наибольшую длину, дан наименьший номер (самыми длинными являются хромосомы 1- и 2-й пары). Выделяют группы метацентрических, субметацентрических и акроцентрических хромосом. Отнесение хромосом к тому или иному типу производится на основе расчета центромерного индекса - отношения длины короткого плеча к длине всей хромосомы. В группе метацентрических хромосом короткое и длинное плечи приблизительно равны, и центромерный индекс приближается к 0, 5. В субметацентрических хромосомах центромерный индекс снижен и составляет от 0, 25 до 0, 35, в акроцентрических хромосомах он часто не превышает 0, 2. На основании комбинации этих двух основных признаков хромосомы сгруппированы в 7 групп, обозначаемых буквами английского алфавита (от А до G).
108. Международная Парижская классификация хромосом человека. Краткий обзор: Латинскими буквами р и q обозначаются соответственно короткое и длинное плечо хромосомы. От центромеры к теломере по имеющимся отчетливым морфологическим указателям (маркерам) в каждом плече выделяют районы, обозначаемые арабскими цифрами. В пределах районов идентифицируют сегменты — регулярные участки, отличающиеся по интенсификации окраски. Они также обозначаются арабскими цифрами. Основная часть: Парижская классификация. В1971 году в Париже на IV международном конгрессе по генетике человека была согласована единая система идентификации хромосом человека, учитывавшая дифференцировку хромосом по длине.Каждая хромосома набора человека при дифференциальной окраске характеризуется уникальным для нее сочетанием темно окрашенных сегментов или полос, чередующихся с неокрашенными участками или светлыми сегментами. Именно такое специфическое для данной хромосомы сочетание сегментов позволяет четко ее идентифицировать и отличить от других хромосом набора. В пределах короткого (р) и длинного (q) плеча каждой хромосомы выделяют ряд четко идентифицируемых областей или регионов, которые нумеруются арабскими цифрами начиная от центромеры к теломерному участку или терминальному концу хромосомы. Каждая область хромосомы включает определенное число сегментов, нумерация которых (второй арабской цифрой) также идет в направлении от центромерного к теломерному участку. Таким образом, обозначение хромосомного сегмента 2q34 означает хромосому №2, длинное плечо, 3 регион и 4 сегмент. Сама центромера обозначается сочетанием цифр 1 и 0, т.е. часть центромеры в пределах короткого плеча обозначается как- р10, а часть, включающая длинное плечо -q10 Открытие в середине 70-х годов того факта, что профазные и про-метафазные хромосомы позволяют достичь большего числа сегментов, чем метафазные хромосомы, и, следовательно, повысить разрешающие возможности цитогенетического исследования, привело к разработке методов получения хромосом высокого разрешения и потребовало дополнения цитогенетической номенклатуры новыми принципами анализа таких хромосом. В1980 году по этому поводу в Париже было достигнуто международное соглашение, которое было опубликовано в 1981 году под названием " Международная система цитогенетической номенклатуры хромосом человека - сегментация хромосом высокого разрешения" или ISCN (1981). Так, если сегмент в пределах какой-либо хромосомы подразделяется на отдельные субсегменты, то после номера сегмента ставится точка, после которой указывается номер субсегмента. Например, если оригинальный сегмент 1 р31 подразделяется на 3 разных субсегмента, то они обозначаются как 1р31.1, 1р31.2и 1р31.3, причем субсегмент 1р31.1 является проксимальным, а 1 р31.3 - дистальным по отношению к центромере. Дополнительное деление субсегментов на другие сегменты, например субсегмента 1 р31.1, соответственно обозначается как 1p31.11, 1р31.12 ит.д.
Вопрос №109 Популярное:
|
Последнее изменение этой страницы: 2016-07-13; Просмотров: 1550; Нарушение авторского права страницы