
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
|
|
Архитектура Аудит Военная наука Иностранные языки Медицина Металлургия Метрология Образование Политология Производство Психология Стандартизация Технологии |
Упругие и сократительные свойства сердечной мышцы
Основной частью сердца является толстостенный левый желудочек, являющийся эллипсоидом вращения, форма которого меняется от почти сферической во время диастолы до сильно вытянутой в конце фазы изоволюмического напряжения (рис 2.15). Мышечные волокна в миокарде расположены в основном циркулярно, и уменьшение их длины (напряжение) приводит к уменьшению окружности миокарда и повышению давления крови в желудочке. Рис 2.15. Изменение размеров и формы полости левого желудочка в разных фазах сердечного цикла: а, б – начало и конец диастолы; в, г – начало и конец систолы Серым цветом отмечено сечение стенки миокарда, черным – объем крови, который находится в левом желудочке Непосредственно перед открытием митрального клапана объем и давление в желудочке минимальны. Согласно закону Лапласа напряжение в миокарде T = F R и поэтому в конце диастолы T увеличивается не менее, чем вдвое, просто из-за увеличения радиуса. Затем наступает фаза изоволюмического напряжения, митральный клапан закрывается, и активно сокращающиеся волокна увеличивают свое напряжение примерно в 25 раз, при некотором уменьшении радиуса, т.к. желудочек становится более вытянутым (принимает форму " огурца" ). Очень важен для понимания физиологии мышечной активности закон Франка – Старлинга: увеличение напряжения в мышце непосредственно перед ее сокращением (преднагрузка) увеличивает силу последующего активного сокращения, или, другими словами: сердце сокращается сильнее во время систолы, если оно в большей степени наполняется во время диастолы.
Рис. 2.16. Иллюстрация закона Франка-Старлинга: 1 – увеличение ударной работы левого желудочка при увеличении среднего давления в левом предсердии 2 – диаграммы " давление-объем" при различной величине МАТЕМАТИЧЕСКИЕ МОДЕЛИ ПРОЦЕССОВ В СИСТЕМЕ КРОВООБРАЩЕНИЯ Упрощенная модель однокамерного сердца Будем считать, что сердечно-сосудистая система организма замкнута и состоит из мышечного (расслабляющегося и сжимающего) однокамерного сердца и одного сосуда, являющейся второй – неактивной, но упругой камерой (рис.3.1). Данная простейшая ситуация имеет некоторый смысл для простейших, червей и моллюсков, но, конечно, никак не для млекопитающих. Рис. 3.1. Эквивалентная электрическая схема системы кровообращения с одной активной камерой
В этом случае кровообращение можно описать линейной системой двух уравнений второго порядка с четырьмя неизвестными: I1 I2
где V1 (t), V2 (t) зависящие от времени и подлежащие определению объемы первой и второй камер, P1 (t), P2 (t) давления в них, F(t) дополнительное внешнее давление, создаваемое активной стенкой первой камеры, а постоянные I1, I2 коэффициенты инерции камер, C1, C2растяжимости, R1, R2 сопротивления камер, тогда как R1, 2 коэффициент сопротивления межкамерного протока. Следует сразу же отметить, что в силу третьего уравнения системы всегда выполнено dV1 (t)/dt = dV2 (t)/dt, т.е. V1 (t) + V2 (t)º V0 º const, (2) что выражает неизменность общего объема крови в двухкамерной системе. Выбирая в качестве внешнего воздействия, которым в системе служит функция напряжения F(t) первой камеры, гармонически изменяющуюся функцию времени F1(t): = F(w) exp(iwt), w - угловая частота, ищем решения системы (1) также в виде гармонических функций с той же угловой частотой вида Vj(t) = где
iwR1, 2 В частности, для статического случая (w = 0) имеем,
P0 = Решение этой системыдается простыми формулами PO =
для гидростатических значений давления в камерах и их объемов. Далее, при w ≠ 0 решение системы (4) для комплексных амплитуд также нетрудно вычисляется:
Имея комплексную амплитуду величины Z(w): = A(w) + iB(w), мы подсчитываем ее модуль |Z(w)|: = Популярное:
|
Последнее изменение этой страницы: 2017-03-08; Просмотров: 462; Нарушение авторского права страницы
 + R1
+ R1  +
+  =
=  , (1)
, (1) + R2
+ R2  +
+  =
=  ,
, 
 (w) exp(iwt), Pj(t) =
(w) exp(iwt), Pj(t) =  (w) exp(iwt), (3)
(w) exp(iwt), (3) j(w),
j(w),  j(w) – комплексные амплитуды, для определения которых непосредственно из (1) получается система линейных алгебраических уравнений
j(w) – комплексные амплитуды, для определения которых непосредственно из (1) получается система линейных алгебраических уравнений (w)(-
(w)(- 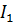 w2 +iwR1 + C1-1) =
w2 +iwR1 + C1-1) =  (w) -
(w) -  (w),
(w),  (w)(-
(w)(-  w2 +iwR2+ C2-1) =
w2 +iwR2+ C2-1) =  (w), (4)
(w), (4) (w)=
(w)=  (w)=
(w)=  (0) + С1
(0) + С1  (0) (5)
(0) (5) (6)
(6)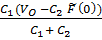 ,
,  ,
, 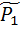 (w) =
(w) =  (7)
(7) (w) =
(w) = 
 (w) =
(w) = 
 (w) = -
(w) = -  , характеризующее максимальное значение, и аргумент arg Z(w) =: j, определяемый равенствами cosj = A/|Z(w)|, sinj = B/|Z(w)| и указывающий сдвиг по фазе относительно опорной гармоники exp(iwt).
, характеризующее максимальное значение, и аргумент arg Z(w) =: j, определяемый равенствами cosj = A/|Z(w)|, sinj = B/|Z(w)| и указывающий сдвиг по фазе относительно опорной гармоники exp(iwt).